Download
1 / 51
720 likes | 1.64k Vues
Експресия на гените. Биологичната информация в ДНК се намира в секвенцията на нуклеотидите във веригата. Генната експресия е процес, при който информацията става достъпна за клетката. Използването на информацията се описва от т. нар. централна догма в молекулярната биология. Генетичен код.

E N D
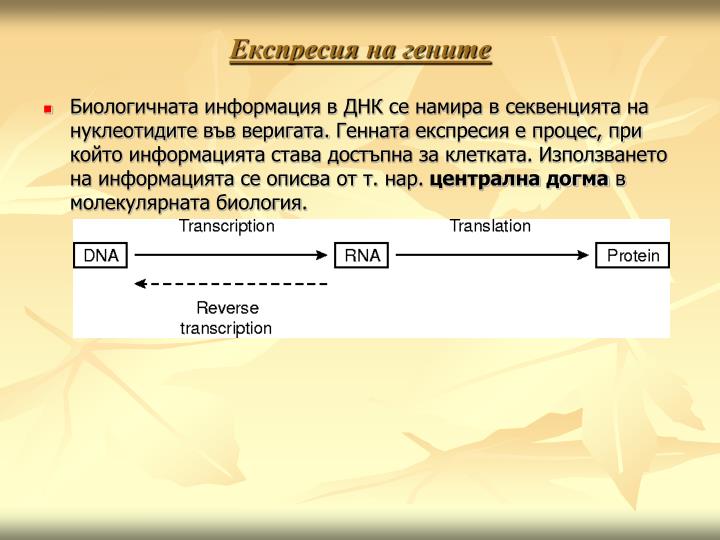
Експресия на гените • Биологичната информация в ДНК се намира в секвенцията на нуклеотидите във веригата. Генната експресия е процес, при който информацията става достъпна за клетката. Използването на информацията се описва от т. нар. централна догма в молекулярната биология.
Генетичен код • Генетичният код описва как секвенциите от нуклеотиди се превръщат в аминокиселиннасеквенция по време на белтъчната синтеза. • Секвенцията на ДНК в гена е разделена на серия от единици, съставени от три бази. Всяка комбинация от три бази се нарича кодон и определя дадена аминокиселина. • Четирите бази в ДНК или в РНК могат да дадат максимална комбинация от 43 = 64 кодона, които определят 20 аминокиселини в белтъка (Таблица 1). • Тъй като броят на кодоните е по-голям, всички АК, с изключение на метионин и триптофан, се кодират от повече от един кодон. • Тази особеност на генетичния код се означава като изроденост
Генетичен код • Кодоните, които определят една и съща АК се наричат синонимни и са сравнително сходни. Например всички кодони ACU, ACC, ACA и ACG определят АК треонин. • Разликите в кодоните са само в базата на трета позиция, която се означава люлееща позиция (wobbling). Изродеността на кода намалява ефекта от мутациите, така че промените в секвенцията на ДНК имат по-малък шанс да променят секвенцията на АК в белтъка и да променят неговата функция. • От всичките 64 кодона 61 са за кодиране на АК. Останалите три, UAG, UGA и UAA не кодират АК, а служат като сигнали за спиране на белтъчната синтеза и са известни като терминиращикодони. • Кодонът за метионин, AUG (GUG), служи като сигнал за начало на транслацията и се нарича иницииращ кодон. • Това означава, че всички белтъци започват с метионин, макар, че понякога тази АК се премахва.
Рамки на четене • Иницииращият кодон не само определя старта на белтъчната синтеза, но дефинира и рамката на четене (readingframe) в секвенцията на РНК. • В зависимост от това коя база е избрана за начало на кодона има три възможни комбинации от кодони, които могат да се прочетат от базовата секвенция. • На практика само една от тези рамки има смислена информация за синтез на белтък; другите две рамки обикновено водят до стоп-кодони, което не позволява използването им за пряка синтеза на белтък (Фиг. 1.11). • Рамка на четене 1: 5’ AUGACUAAGAGAUCC GG -3’ • MetThrLysArgSer • Рамка на четене 2: 5’A UGACUAAGAGAUCCG G -3’ • StopLeuArgAspPro • Рамка на четене 3: 5’ AU GACUAAGAGAUCCGG -3’ • AspStopGluIleArg • Всяка секвенция ДНК може да бъде четена в три отделни рамки, в зависимост от това от коя база започва кодона.
Транспортна РНК • Роля в транслацията • Клетките съдържат три вида РНК – транспортна, рибозомна и информационна; транскрибират се от ДНК. • Транспортната и рибозомната РНК са част от апарата на белтъчната синтеза, а иРНК участва като матрица за синтеза на белтъци по време на транслацията. • Транспортните РНК (тРНК) са малки молекули, които действат като адаптери по време на белтъчната синтеза. • Те са връзката между нуклеотиднатасеквенция на иРНК и аминокиселиннатасеквенция на белтъка. • Клетките съдържат определен брой тРНК, всяка от които може да се свързва само с определена аминокиселина (АК). • Всяка тРНК различава един кодон в иРНК, което ú позволява да поставя АК на правилна позиция в растящата полипептидна верига, както е определено от секвенцията на иРНК.
Транспортна РНК - структура Акцепторно рамо. Секвенцията ССА, която се намира на 3’-края на веригата на всички тРНК е несдвоена с други нуклеотиди и нуклеотида А е мястото на свързване на аминокиселината. D-рамо (или DHU) е структура стъбло-бримка, съдържащо дихидроурацил (D) нуклеотид с модифицирана азотна база. Антикодоново рамо, отговорно за разпознаването и свързването на кодона в иРНК. Комплементарен на кодона. Допълнително или вариабилно рамо. Може да е малко, съдържащо само 2-3 нуклеотида (клас I тРНК) или по-голямо, съдържащо 13-21 нуклеотида (клас II тРНК). ТψС рамо - съдържа секвенцията ТψС, където с ψ (пси) се означава необичайният, модифициран нуклеотид псевдоурацил.
Транспортна РНК – 3D структура – L - форма • Детелиновият лист е двуизмернатаформана тРНК. • Триизмерната структура (3D) е получена от данни от рентгено-структурен анализ (фигурата). • Нуклеотидите, свързани в 2-D структура са свързани и с некомплементарни водородни връзки между нуклеотиди, които са раздалечени и са на различни рамене като те образуват 3D структурата.
Транспортна РНК – 3D структура – L - форма • Тези връзки се наричат третични, тъй като образуват третичната структура на тРНК. • Много от тези нуклеотиди са консервативниили полу-инвариантни. • В 3D структурата акцепторното рамо и антикодоновото рамо са на противоположнитекраища на молекулата в съответствие с различната им функции в транслацията
Рибозоми • Рибозомите са макромолекулни структури, изградени основно от белтъци и рибозомна РНК(рРНК), свързани в надмолекулен комплекс. • Рибозомите се намират в клетъчната цитоплазма. • Клетъчните нужди за синтез на белтък изискват голям брой рибозоми. • В една типична бактериална клетка има около 20 000 рибозоми, които представляват около 80% от цялата РНК в клетката и около 10% от клетъчните белтъци. • Тъй като рибозомите са много големи е трудно да се получат данни за тяхната молекулна маса. • Вместо това рибозомите се измерват по тяхната константа S, известна като седиментационна константа (константа на Сведберг), която отразява движението на подобни комплекси във вискозни среди (разтвор на захароза или на цезиев хлорид), което става при центрофугиране. • Стойността S се определя от големината, формата и макро-молекулната структура на рибозомата.
Рибозоми • Всяка рибозома се състои от две части, наречени голяма и малка субединици. • В прокариоти като E. coli рибозомата е 70S и е съставена от две субединици – 50S и 30S (както се вижда S стойностите не се събират; не са адитивни). • Голямата субединица (50S) съдържа две рРНК (23S и 5S) в комплекс с 31 рибозомни белтъка. • 30S субединицата съдържа една единствена рРНК (16S) и 21 белтъка. • Еукариотите рибозомите са 80S и се състоят от 60S и 40S субединици. • Голямата 60S субединица съдържа три рРНК – 28S, 5.8S и 5S и около 40 белтъка; • 40S субединицата съдържа една единствена рРНК (18S) и около 33 белтъка.
Информационна РНК. Процесинг, сплайсинг • При еукариотите информационната РНК (иРНК) се синтезира при транскрипцията на гени, кодиращи синтезата на белтъци, от ензима РНК полимераза II. • Тя служи за матрица за синтез на белтък при транслацията. • Кодиращата информация на такива гени е фрагментирана и е организирана като поредица от екзони, разделени от некодиращи интрони. • и РНК се синтезира като предшественик, наречен пре-иРНК, която е копие и на екзоните и на интроните. • Преди да послужи като матрица за синтез на белтък, пре-иРНК преминава през серия процеси на зреене (processing), за да се получи зрялата иРНК. • Некодиращите секвенции на интроните се отделят с процеса сплайсинг (splicing), който свързва кодиращите секвенции и осигурява функционална иРНК за транслация.
Информационна РНК. Кепиране и полиаденилиране • В допълнение 5’-края се модифицира чрез добавяне на един модифициран нуклеотид, процес, известен като кепиране (caping). • От своя страна 3’-края се модифицира чрез добавяне на около 250 аденинови нуклеотида. • Процесът е известен като полиаденилиране. • Молекулите РНК, синтезирани от ензима РНК полимераза II съществуват като популация от молекули с различна дължина и на различни етапи на зреене се означават като хетерогенна ядрена РНК или хяРНК (hnRNA). • В прокариотните иРНК не се процесира и транслацията започва на 5’ края дори още преди молекулата да е синтезирана изцяло. • Нормално прокариотните гени не съдържат интрони и затова сплайсинг не е необходим.
Кепиране • ЕукариотнитеиРНК се модифицират на своя 5’-край с един процес, известен като кепиране(capping), който включва добавяне на един модифициран нуклеотид, 7-метилгуанозин. • Той се добавя с ензима 7-метилгуанозин трансфераза, който свързва GTP с необичайната 5’-5’ фосфодиестер-на връзка. • След това един ензим, метилтрансферазадобавя метилова група при азота на седмо място в гуаниновия пръстен, както и към 2’-ОН групите на следващите два нуклеотида. • Кепирането предотвратява разграждането на 5’-края на иРНК от екзонуклеазите на цитоплазмата, а също е сигнал за рибозоматаза разпознаване на началото на транслацията.
Полиаденилиране • Повечето еукариотни пре-иРНК са модифицирани на своя 3’-край с добавяне на секвенция от 250 нуклеотида - поли-А опашка. • Тази модификация се нарича полиаденилиране и изисква наличието на сигнална секвенция в пре-иРНК. • Тя се нарича сигнална секвенция за полиаденилиране, 5’AAUAAA 3’, и се намира близо до 3’-края на пре-иРНК. • В следващите 11-20 бази се намира секвенция YA (Y – пиримидин), а по-нататък се намира друга секвенция, богата на GC нуклеотиди. • Няколко специфични белтъка разпознават и се свързват с тези сигнални секвенции и образуват комплекс, който реже иРНК на няколко нуклеотида след секвенцията 5’AAUAAA 3’. • След това ензимът поли-А полимераза добавя аденинови нуклеотиди към 3’-края. • Ролята на поли-А опашката не е добре известна, но може да предпазва 3’-края на кодиращата секвенция от разграждане от цитоплазмените рибонуклеази. • Въпреки това са известни случаи, като особено типични са иРНК за хистоните, където няма поли-А опашка.
Транслация. Роля на тРНК • Транслацията е процес, чрез който клетките синтезират белтък. • По време на транслацията, информацията, кодирана от иРНК се използва, за да определи секвенцията на АК в белтъка. • Транспортната РНК играе ключова роля в този процес, чрез осигуряване и транспортиране на АК до рибозомата в ред, който е определен от иРНК. • Това гарантира, че АК ще се свържат в правилна последователност
Транслация. Роля на тРНК • Клетките съдържат от 31 до 40 отделни тРНК, всяка от които се свързва с една от 20-те АК. • Съответно може да има повече от една тРНК за дадена АК. • Транспортни РНК, които свързват една АК се означават като изоакцепторни. • Преди да започне белтъчната синтеза АК се свързват ковалентно с тРНК, които след това разпознават кодона в иРНК, отговарящ на дадена АК.
Транслация. Аминоацилиране • Свързването на АК към тРНК се нарича аминоацилиранеили зареждане. • АК се свързва ковалентно с края на акцепторното рамо на тРНК, което винаги завършва със секвенцията 5’CCA 3’. • При това се формира връзка между 3’-ОН групата на крайния аденин (А) на акцепторното рамо и карбоксилната група на аминокиселината. • Зареждането се катализира от ензим, наречен аминоацил-тРНК синтетаза с реакция, която изисква хидролиза на АТФ (АТР).
Транслация. Аминоацилиране • За всяка АК има отделен ензим и този ензим може да зареди всички изоакцепторни тРНК, отговарящи за тази АК. • Ензимът аминоацил-тРНК синтетаза разпознава едновре-менно АК и съответната тРНК. • АК се разпознава основно по страничната си верига. • Не е много ясно как се разпоз-нава тРНК, но се предполага, че причина са различни нуклеоти-ди, специфични за всяка тРНК.
Транслация. Инициация • Когато не са активно използвани в транслацията, рибозомите са като отделниголеми и малки субединици. • Първата стъпка на транслацията представлява свързване на малката субединица към иРНК (Фигура). • Транслацията започва обикновено при кодона АUG (при бактериите GUG и UUG), който кодира АК метионин и е известен кат иницииращ кодон. • Малката субединица на рибозомата (30S) се свързва в определен участък преди AUG (upstream). • При прокариотите това се означава като секвенцияШайн-Далгарно (Shine-Dalgarno, 5’ AGGAGGU 3’), която е близо до старта на иРНК.
Транслация. Инициация • Веднъж свързана, 30S мигрира в посока 3’ по иРНК, докато не намери AUG, което е обикновено на 10 нуклеотида по-надолу (downstream). • При еукариотите малката рибозомна субединица разпознава кепирания 5’-край на иРНК. • След това тя се движи по иРНК, дока-то не разпознае първия AUG кодон, • Към AUG кодона, локализиран в малката субединица се свързва тРНК, заредена с метионин (тРНКmet). • При бактериите метионинът е модифициран чрез свързване на една формилна група (-СНО) към водородния атом на аминогрупата (тРНКfmet).
Транслация. Инициация • Комбинацията от тРНКfmet, иРНК и 30S се нарича иницииращ комплекс • За разпознаване на AUG има двеmet-тРНК – за инициация (тРНКifmet)и за вътрешните AUG кодони. • Само инициаторната тРНК е може да се свърже в инициаторния комплекс. • За инициацията са необходими допълнителни белтъци, наречени иницииращи фактори (IF). • Бактериите имат три, известни като IF1, IF2 и IF3. • Инициацията започвачрез свързване на IF1 и IF3 към 30S. • След това 30S се свързва с иРНК и намира иницииращия AUG кодон • Към комплекса се свързва ин-тРНК, заредена с метионин и се отделя IF3. • Това e краятна инициацията.
Еукариотна инициация на транслацията • Инициацията при еукариотите е сходна с бактериалната, с тази разлика, че иницииращия кодон тук е главно AUG, а метионинът не е формилиран. • Тук участват значително повече фактори (поне 9), като някои от тях са хомоложни с бактериалните. • Има два фактора (eIF2 I eIF 3), чиято функция е сходна с техните аналози при бактериите (IF2 и IF3). • Няколко от еукариотните фактори са отговорни за премахване на вторични структури по иРНК, преди тя да бъде транслирана. • Енергия за това се взема от хидролизата на ATP. • Един специфичен еукариотен фактор е eIF-4B, който участва в разпознаването на кепирания 5’ край на еукариотната иРНК.
Транслация. Елонгация • Веднага след формирането на иницииращия комплекс, към него се свързва 50S. • Рибозомата има два свързващи участъка (сайта) за тРНК молекули (Фиг. 1.28). • Първият участък е Р или пептидиленсайт и е зает от тРНКifmet, свързана с кодона AUG. • Вторият участък е А или аминоацилен сайт и е разположен върху втория кодон. • Елонгацията започвакогато заредена тРНК навлезе в А и се свърже с втория кодон. • В този момент в рибозомата има две заредени тРНК, а двете им АК са в близо и между тях се образува пептидна връзка. • Реакцията за образуване на пептидната връзка се осъществява от голямата рРНК на 50S, която в случая играе ролята на рибозим.
Транслация. Елонгация • За елонгацията са необходими елонга-ционнифактори (EF). • При бактериите има два фактора, EF-Tu и EF-Ts за навлизането на тРНК в А-участъка. • ЕF-Tu се свързва с тРНК, като предварително се е свързал с GTP. • След навлизането в А участъка GTP се хидролизира и от рибозомата излиза EF-Tu, свързан с GDP. • Преди да се свърже с друга АК, EF-Tu се регенерира с помощта на EF-Ts. • Първоначално EF-Ts измества GDP, а след това нова молекула GTP се свързва с EF-Tu, измествайки EF-Ts (фигурата по долу).
Транслация. Елонгация • След образуването на пептидната връзка се наблюдава транслокация, което е придвижване на иРНК на следващия кодон. • Формираният дипептид (аа-аа) се придвижва в Р участъка, измествайки вече празната тРНК, която се намира там. • При това А участъкът остава свободен и в него може да навлезе нова тРНК, при което елонгационният цикъл се повтаря. • При бактериите в транслокацията се участва EF-G или транслоказа, който се свързва в рибозомата, свързан с GTP. • И тук се извършва хидролиза на GTP, при което се отделя енергия за транслокацията. • Свързването на EF-Tu и на EF-G е алтернативнои взаимно-изключващо се. • Това гарантира, че транслокацията ще се извърши преди да започне нов цикъл на елонгация.
Транслация. Терминация • Транслацията завършва, когато в А участъка попадне терминационен кодон (Фиг. 1.30). • Не съществуват тРНК, които да се свързват с терминационния кодон. • Вместо това в А участъка навлизат RF - терминиращи фактори (release factors), които предизвикват отделянето на завършения белтък. • В E. coli този процес се изпълнява от RF-1 и RF-2. • RF-1 разпознава UAA и UAG, докато RF-2 разпознава UAA и UGA. • Трети терминиращ фактор RF-3 играе допълнителна роля в процеса. • Тези фактори прехвърлят последната АК на формирания белтък върху молекула вода, вместо към следващата аминоацил тРНК.
Репликация на ДНК • Това е процес, при който в клетката се копира ДНК. • Репликацията е необходима за предаване на генетичната информация в клетката да бъде предавана на дъщерните клеткипо време на клетъчното делене. • Репликацията се означава като полуконсервативен процес (Фиг. 1.31). Това означава, че новополучената ДНК има една стара верига, получената от майчината ДНК и една дъщерна, новосинтезирана верига.
Репликация на ДНК • Механизмите на репликация са много сходни при много различни организми. • Разликите засягат основно ензимите на репликация и участващите белтъци. • В прокариотните организми като E. coli за репликацията отговарят два ензима – ДНК полимераза I и ДНК полимераза III. • При еукариотите ДНК се реплицира от 5 ензима (ДНК полимераза α, β, γ, δ и ε).
Репликация на ДНК • Репликацията трябва да е много точен процес, защото дори много ниско ниво на грешки може да доведе до сериозни промени само след няколко репликации. • Точността на репликацията се гарантира от способността на ензима да проверява дали при удвояването е включен правилният нуклеотид. • Това става с обратна 3’-5’ екзонуклеазна активност на ензима, която му позволява да отдели неправилно присъединен нуклеотид и след това да го замести с правилен, използвайки 5’-3’ полимеразна активност. • Това се означава като коригиране или репарационно свойство на ДНК полимеразата. • Критичния минимум на грешки при репликацията е една грешка на 5 милиарда нуклеотида.
Репликативна вилка • Разплитането на двойната верига започва от определено място, наречено начало на репликацията (origin) • Мястото, където се разплита ДНК и се синтезират новите вериги се наричарепликативна вилка
Синтеза на водещата и закъсняващата вериги • Синтезата на ДНК от ДНК полимеразата става само в посока 5’-3’. • Тъй като двете вериги на ДНК са антипаралелни (едната има посока 5’-3’, а другата – 3’-5’) за тяхната репликация се използват малко различаващи се механизми. • Едната верига, наречена водеща (leading) се реплицира в същата посока, в която се развива веригата, така че синтезата може да става непрекъснато. • Другата верига, известна като закъсняваща (lagging) се синтезира в противоположна посока и трябва да се синтезира прекъснато. • Тя се синтезира като серия от сегменти, наречени фрагменти на Оказаки
Синтеза на водещата и закъсняващата вериги
Синтеза на водещата и закъсняващата вериги
Начало на репликацията (priming). • ДНК полимеразата изисква къс, двойноверижен участък, за да инициира синтезата на ДНК. • Този участък се нарича праймер (primer) и се образува от ДНК полимераза, наречена праймаза, която има способността да започне синтезата върху единична верига ДНК. • Праймазата синтезира къс РНК праймер върху матрицата, при което се получава къс едноверижен участък. • При E. coli след това ДНК полимераза III синтезира ДНК, започвайки от праймера.
Начало на репликацията (priming). • При закъсняващата верига синтезата спира, когато се стигне до следващия праймер. • В този момент започва да действа ДНК полимераза I, която премахва РНК праймера и го замества с ДНК (Фиг. 1.34). • При еукариотите ситуацията е различна. Там ДНК полимераза α, която има цялостна праймазна активност е отговорна за инициация на репликацията. • ДНК се реплицира от ДНК полимерза α и δ, като δ синтезира водещата верига, а α - закъсняващата. • Другите полимерази имат допълваща роля. ДНК полимераза ε участва в репарацията на ДНК, а РНК полимераза γ реплицира ДНК при митохондриите.
Репликация на ДНК при бактериите • За репликация на пръстеновидните ДНК на бактериите и на линейните ДНК на еукариотите се прилагат различни стратегии. • Използва се едно начало на репликация, от което започват две репликативни вилки, движещи се в противоположни направления. • При това се получават междинни репликативни форми, напомнящи гръцката буква θ (тита), което дава и името на модела (Фиг. 1.35). • В края на краищата двете вилки се срещат, сливат се и с това репликацията завършва.
Репликация на ДНК при еукариотите • Преди клетката да се раздели в нея трябва да се реплицира ДНК. • Клетъчното делене при еукариотите е във висока степен регулирано и става на серия от различни фази, означавани като клетъчен цикъл (Фиг. 1.36). • Продължителността на цикъла варира, но обикновено е няколко часа. • Най-дългата фаза е G1, при която клетката се приготвя за делене. • Тя е последван от S фазата, в която става репликация на ДНК. Следва по-къс G2 период, последван от фаза М. • През този период клетката преминава митоза, през която разделяне на хромозомите и делене на клетката. • След митозата делящите се клетки навлизат в G1 фазата на следващия клетъчния цикъл. В други случаи клетката може да излезе от клетъчния цикъл, навлизайки в периода G0, в който не се делят за определено време. Нервните клетки са винаги в период G0.
Регулация на генната експресия – обща характеристика • За да функционират бактериите не е необходимо всички гени да бъдат транскрибирани едновременно. • За да се запази енергията и ресурсите бактерията регулира активността на гените си, така че се експресиратсамо тези гени, чиито продукти са необходими в дадения момент. • Например ще бъде излишен разход за бактерията да произвежда ензими, които са необходими за синтез на аминокиселина, която в момента е налична в клетката. • Регулацията на генната експресия позволява бактерията да отговаря на промените на външната среда, най-често по отношение на наличие или липса на хранителни вещества.
Организация на бактериалните гени • Една важна характеристика, отразяваща се на начина, по който се регулират гените, е тяхната организация в оперони. • Това са транскрипционни единици от няколко гена, кодиращи белтъци със сродна функция, които се регулират заедно. • Има и други гени, кодиращи регулаторни белтъци, които контролират генната експресия на опероните. • В E. coli са установени много различни оперони. Най-много са опероните, кодиращи ензими за синтеза на аминокиселини или за разграждане на различни субстрати. • Опероните се класифицират на индуцибилни и репресибилни.
Организация на бактериалните гени • Индуцибилнитеоперони съдържат гени, които кодират ензими, участващи в метаболитните пътища. • Експресията на гените се контролира от субстрата на този метаболитен път. • Пример за индуцибиленоперон е лактозния (lac), кодиращ ензими, извършващи разграждането на лактозата. • Репресибилнитеоперони съдържат гени, които кодират ензими на биосинтетични пътища и експресията се регулира от крайния продукт на тези пътища, които могат да репресират експресията на оперона или да я контролира чрез алтернативен механизъм атенюация. • Пример за репресибиленоперон е триптофановия (trp), който кодира ензими за синтез на триптофан.
Лактозен (lac) operon • Оперонът съдържа три гена, кодиращи ензими за утилазацията на дизахарида лактоза в клетката на E. coli. • Това са β-галактозид-пермеаза, който транспортира лактозата в клетката, β-галактозидаза, който хидролизира лактозата на два монозахарида (глюкоза и галактоза) и β-галактозидтрансацетилаза, който ацилира лактозата и също участва в хидролизата на лактозата. • Тези ензими се съдържат в клетката в много малка концентрация, но в присъствие на лактоза нивото им бързо нараства. • Трите гена са известни като lac A, Z и Y. Те са подредени последователно и се транскрибират в една единствена иРНК от един промотор
Лактозен (lac) operon • Преди оперона (upstream) се намира регулаторен ген, lac I, който се експресира и кодира синтезата на белтък, наречен lacрепресор, който регулира експресията на гените lac Z, Y и A. • При липса на лактоза репресорът се свързва с секвенция ДНК, наречена оператор, който се намира между промотора и първия ген lac Z. • Когато е свързан с оператора, репресорът блокира пътя на РНК полимеразата за свързване с промотора и предотвратява транскрипцията. • Когато клетката се срещне с лактоза тя се свързва с лактозниярепресор и променя конформацията му така, че той повече не може да се свързва с оператора.
Катаболитна репресия • Този термин означава един допълнителен регулаторен механизъм, който позволява на lacоперона да отчита наличието на глюкоза, която е алтернативен и предпочитан въглероден енергиен източник от бактерията. • Ако в клетката има глюкоза и лактоза, клетката ще използва първо глюкозата с предимство, вместо да използва енергия да разцепва лактозата и тогава да използва глюкозата от този процес. • Присъствието на глюкоза изключва lacоперона чрез един механизъм, известен като катаболитна репресия, който използва регулаторен белтък, наречен САР (cataboliteactivatorprotein)
Катаболитна репресия • САР се свързва с оперона в област преди лактознияпромотор и ускорява свързването на РНК полимеразата към промотора. • При това обаче САР действа само когато е свързан с цикличен аденозинмонофосфат (cAMP), чието ниво се определя от нивото на глюкозата. • Ензим, нареченаденилатциклазасинтезира cAMPoт ATP и се потиска от глюкозата. Когато в клетката има глюкоза, аденилатциклазата се инхибира и нивото на cAMP е ниско. • При тези условия САР не се свързва с промотора и lacоперона се експресира слабо. Обратно, когато глюкозата е малко, нивото на cAMP е високо, САР се свързва и нивото на експресия на lacоперона е високо. • Ако има едновременно лактоза и глюкоза, lacоперона ще се експресира при много ниско ниво. Когато обаче глюкозата се разгради, катаболитната репресия се премахва и lacоперонът се експресира в зависимост от нивото на лактозата
Триптофанов (trp) оперон • Оперонът съдържа пет гена, кодиращи ензими, участващи в биосинтезата на аминокиселината триптофан. • Гените се транскрибират в единична иРНК, транскрибирана от един промотор. • Експресията се регулира от нивото на триптофана в клетката. Един регулаторен ген, намиращ се преди промотора синтезира trpрепресор. • Той се свързва с trp оператор, който се намира непосредствено след trpпромотор, като дори частично се припокрива с него. • Когато в клетката има триптофан, той се свързва с репресора и го активира за свързване с оператора, пречейки на свързването на РНК полимеразата с промотора и по този начин спирайки транскрипцията на оперона. • При липса на триптофанрепресорът не може да се свърже с оператора и оперонът е свободен да се транскрибира. • По този начин триптофанът, като краен продукт от синтезата служи като ко-репресор, заедно с trpрепресор и инхибира собствената си синтеза.
