Download

1 / 43
500 likes | 731 Vues
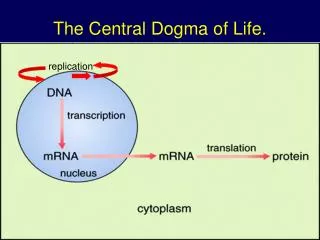
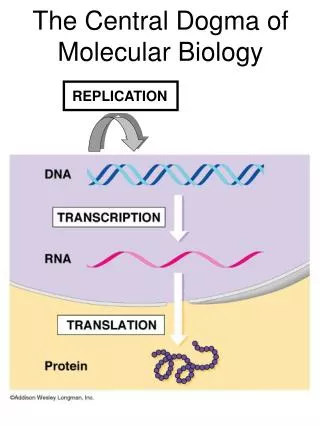
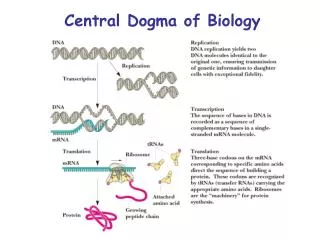
MCB 110:Biochemistry of the Central Dogma of MB. Part 1. DNA replication, repair and genomics (Prof. Alber). Part 2. RNA & protein synthesis. Prof. Zhou. Part 3. Membranes, protein secretion, trafficking and signaling. Prof. Nogales. MCB 110:Biochemistry of the Central Dogma of MB.

E N D
MCB 110:Biochemistry of the Central Dogma of MB Part 1. DNA replication, repair and genomics (Prof. Alber) Part 2. RNA & protein synthesis. Prof. Zhou Part 3. Membranes, protein secretion, trafficking and signaling Prof. Nogales
MCB 110:Biochemistry of the Central Dogma of MB Part 1. DNA replication, repair and genomics (Prof. Alber) Part 2. RNA & protein synthesis. Prof. Zhou Part 3. Membranes, protein secretion, trafficking and signaling Prof. Nogales
DNA structure summary 1 W & C (1953) modeled average DNA (independent of sequence) as an: anti-parallel, right-handed, double helixwithH-bonded base pairs on the inside and the sugar-phosphate backbone on the outside. Each chain runs 5’ to 3’ (by convention). Profound implications: complementary strands suggested mechanisms of replication, heredity and recognition. Missing Structural variation in DNA as a function of sequence Tools to manipulate and analyze DNA (basis for biotechnology, sequencing, genome analysis)
DNA schematic (no chemistry) 1. Nucleotide = sugar-phosphate + base DNA strands are directional 3. Duplex strands are antiparallel and complementary. Backbone outside; H-bonded bases stacked inside. 4. The strands form a double helix
Nucleic-acid building blocks nucleoside glycosidic bond nucleotide
C G T A Geometry of DNA bases and base pairs! H-bonds satisfied Similar width Similar angle to glycosidic bonds Pseudo-symmetry of 180° rotation
Major groove Major groove Major groove and minor groove definitions Opposite the glycosydic bonds Minor groove Minor groove Subtended by the glycosydic bonds
3.4- 3.6 Å Comparison of B DNA and A DNA (formed at different humidity) Bps near helix axis Bps off helix axis Major groove (winds around) Minor groove (winds around) bp/turn Base tilt Major groove Minor groove P-P distance 10 small wide Narrow 6.9 Å 11 20° narrow & deep wide & shallow 5.9 Å
Average structure of dsRNA (like A DNA) 3’ 5’ Minor groove shallow and wide Major groove deep and narrow (distortions needed for proteins to contact bases) “side” view Bases tilted 5’ 3’ Twist/bp ~32.7° ~11 bp/turn “End” view
DNA structure and stability DNA structure varies with sequence 1. “Dickerson dodecamer” crystal structure 2. Twist, roll, propeller twist and displacement 3. Variation in B-DNA and A-DNA Proteins recognize variations in DNA structure DNA stability Depends on sequence & conditions Forces that stabilize DNA: H-bonds, “stacking”, and interactions with ions and water
Crystal structure of the “Dickerson dodecamer” Experiment -- 1981 Synthesize and purify 12-mer: d(CGCGAATTCGCG) = sequence Crystallize Shine X-ray beam through crystal from all angles Record X-ray scattering patterns Calculate electron density distribution Build model into e- density and optimize fit to predict the data Display and analyze model Results B-DNA!! The structure was not a straight regular rod. There were sequence-dependent variations (that could be read out by proteins).
Two views of the Dickerson dodecamer Double helix: Anti-parallel strands, bps “stacked” in the middle Not straight (19° bend/12 bp, 112 Å radius of curvature) Core GAATTC: B-like with 9.8 bp/turn Flanking CGCG more complex, but P-P distance = 6.7 Å (like B) Bps not flat. Propeller twist 11° for GC and 17° for AT Hydration: water, water everywhere on the outside (not shown).
Nomenclature for helical parameters Propeller twist: dihedral angle of base planes. Displacement: distance from helix axis to bp center Slide: Translation along the C6-C8 line Slide Twist: relative rotation around helix axis Roll: rotation angle of mean bp plane around C6-C8 line Tilt: rotation of bp plane around pseudo-dyad perpendicular to twist and roll axes
Propeller twist, roll and slide Slide = -1 Å to avoid clash * No roll or propeller twist 20° propeller twist Or roll = 20 ° and slide = + 2Å to promote cross-chain purine stacking
Slide and helical twist Slide = translation along the long (C6-C8) axis of the base pair
Regular DNA variations B-like A-like
Helical parameters of the dodecamer C1/G24 G12/C13 Range 4.9-18.6° 32.2-41.4° 8.1-11.2 3.14-3.54 Å
Helical parameters of the dodecamer C1/G24 G12/C13 Range 4.9-18.6° 32.2-41.4° 8.1-11.2 3.14-3.54 Å
Helical parameters of the dodecamer C1/G24 G12/C13 Range 4.9-18.6° 32.2-41.4° 8.1-11.2 3.14-3.54 Å
Base “stacking” maximizes favorable interactions Clashes due to propeller twist can be alleviated by positive roll (bottom left) or changes in helical twist (right) N atoms close N atoms separated roll helical twist
Different patterns of H-bond donors and acceptors bases in different base pairs (gray) Major groove side (w) Most differences in H-bond donors and acceptors occur in the major groove! Sequence-specific recognition uses major-groove contacts. Seeman, Rosenberg & Rich (1976), Proc Natl Acad Sci USA73, 804-8. Minor groove side (S)
Lac repressor headpiece binds differently to specific and nonspecific DNAs Symmetric operator Natural operator Bent DNA Straight DNA Nonspecific DNA
E. coli lac repressor tetramer binds 2 duplexes Headpiece Hinge helix NH2 N-subdomain C-subdomain Tetramerization helix LacI tetramer
E. coli lac repressor tetramer binds 2 duplexes Headpiece Hinge helix NH2 N-subdomain C-subdomain Tetramerization helix Repressor tetramer loops DNA
E. coli catabolite activator protein (CAP) Stabilizes kinks in the DNA
Human TATA binding protein binds in the minor groove and stabilizes large bends Twist along the DNA DNA bent
Human TATA binding protein binds in the minor groove and stabilizes large bends TBP TBP DNA View into the saddle End view
DNA bending by E. coli AlkA DNA glycosylase Leu125 inserted into the DNA duplex! 66° bend
Base flipping in DNA repair enzymes Human Alkyl Adenine DNA Glycosylase Phage T4 A Glycosyl Transferase,AGT
What cause bases to flip out? Thermal fluctuations
Fluctuations include denaturation Native Denatured + Tm = 50/50 native/denatured T
Tm depends on? DNA LengthBase composition DNA Sequence Salt concentration Hydrophobic and charged solutes Bound proteins Supercoiling density
Length dependence of DNA stability No further increase > ~50 base pairs 10 20 30 Fraction denatured Temperature °C
Tm depends on G+C content Why? GC bps contain 3 H-bonds and stack better.
Calculated base stacking energies GC best AT worst
Tm depends on ionic strength High KCl stabilizes duplex DNA Why?
Other conditions that change Tm } Mg2+ ions Polyamines: spermidine and spermine + + + NH3-CH2-CH2-CH2-NH2-CH2-CH2-CH2-CH2-NH3 NH3-CH2-CH2-CH2-NH2-CH2-CH2-CH2-CH2-NH2-CH2CH2-CH2-NH3 + + + + DMSO formamide H3C CH3 HC NH2 C Stabilize (why?) } Destabilize (why?) O O
Duplex stability depends on length (to a point) and base composition (GC content) Two formulas for oligonucleotide Tm 1. Tm = (# of A+T) x 2 + (# of G+C) x 4 2. Tm= 64.9 +41 x ((yG+zC-16.4)/ (wA+xT+yG+zC)) where w, x, y, z are the numbers of the respective nucleotides.
Summary DNA structure varies with sequence. Propeller twist, helix twist, roll, slide, and displacement (local features) vary in each base step. These differences alter the positions of interacting groups relative to ideal DNA. Structural adjustments maximize stacking. Proteins can read out base sequence directly and indirectly (e.g. H2O, PO4 positions, structure and motions). Proteins can trap transient structures of DNA. Duplex stability varies with sequence, G+C > A+T High salt, Mg2+, polyamines increase duplex stability. DMSO and formamide decrease duplex stability. Stability increases with oligonucleotide length up to a point.