Download

1 / 32
350 likes | 593 Vues
Programmed Cell Death A genetically controlled cell suicide pathway. The History of the Cell Death Study. First observed in 1842 by a German scientist Carl Vogt in Jent und Gassman, Solothurn 1842; 130

E N D
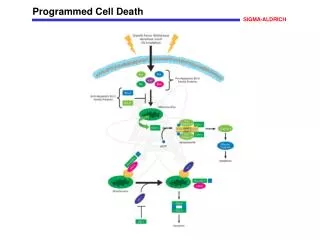
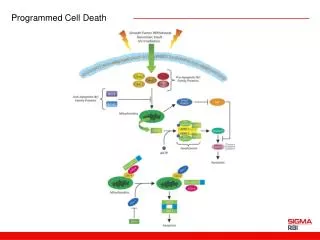
Programmed Cell Death A genetically controlled cell suicide pathway
The History of the Cell Death Study • First observed in 1842 by a German scientist Carl Vogt in Jent und Gassman, Solothurn 1842; 130 He wrote “The role of the core of the notochord in the formation of the vertebrae is quite simply that its cells are resolved , beginning when the proliferation of the surrounding cartilage exerts pressure on the notochore”. “The light vesicular nuclei of the embryonic cells have disappeared; at least I could not detect any trace of them”. • 1951, Glucksmann, A (in Cambridge Philosophical Society of bilogical review 26, 59) reviewed and rediscovered developmental cell deaths in embryological tissues • 1964, Lockshin, RA and Williams, CM first used the term of “programmed cell death” to describe the breakdown of the intersegmental muscles of silkworms • 1972, three British scientists Kerr, Wyllie and Currie proposed the term of “apoptosis” for morphology of naturally occurring or physiological cell deaths. They recognized the importance of apoptosis and suggested that it is a basic biological phenomenon with wide-ranging implications in tissue kinetics.
Chromosomes condense and fragment Nuclear membrane breaks down Apoptotic body formation Engulfment of the cell corpse The Morphology of Apoptosis Cytoplasm shrinks
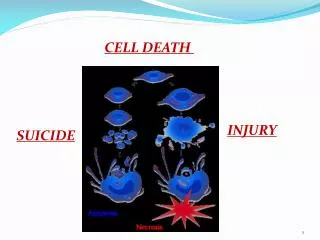
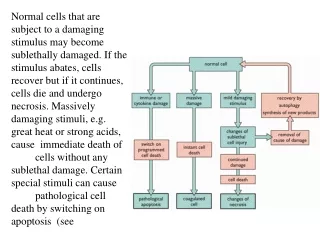
Difference Between Apoptosis and Necrosis • Necrosis (pathological cell death): dying cells swell and lyse; toxic contents leak out and result in inflammatory response. • Apoptosis (physiological or programmed cell death): dying cells shrink, are engulfed by other cells, leave no trace, and don’t result in harmful outcomes
Serve some function but no longer needed e.g. tadpole tail of frog. Needed in one sex but not another e.g. Mullerian duct important for female is eliminated in males by apoptosis. Serve in immune system as a defense mechanism to get rid of harmful or damaged cells. Functions of apoptosis Sculpt body structures, e.g. hand digit Produced in excess, e.g. extra neurons are removed by apoptosis during neurogenesis.
The Nematode C. elegans As a Model Organism in the Study of PCD • A great genetic system • Completely defined cell lineage • Study of cell death at a single cell resolution in living animals
The C. elegans Cell Lineage z y g o t e A B M S E C D 4 P
Cell Death Can Be Studied at a Single Cell Resolution P11 X X P11aap Adapted from Sulston and Horvitz, Dev. Bology 56, 110-150, 1977
Wild-type nuc-1 The First Few Cell Death Mutants Identified in C. elegans In 1976, J. Sulston first described programmed cell death in nematodes and reported the first cell death mutant (nuc-1), in which DNA in the death cells fail to be degraded.
Wild-type ced-1 In 1980, E. Hedgecock isolated two cell death mutants (ced-1 and ced-2) which are pivotal for identification of the other cell death genes. What went wrong with the ced-1 mutant?
Phenotypic analysis of ced-1 and ced-2 mutants More cell deaths? Dying cells cannot be removed or engulfed How to distinguish these two possibilities? Follow the cell lineage in the mutant animals What is next?
Suppressor screens:ced-3 and ced-4 What are the functions of ced-3 and ced-4? H. Ellis and R.H. Horvitz
What are the functions of ced-3 and ced-4? ced-3 and ced-4 promote cell corpse engulfment? Then the mutations must be increase-of-function 2) Inhibitors of cell corpse engulfment? Then the mutations should be loss-of-function 3) ced-3 and ced-4 could promote cell deaths Then the mutations should be loss-of-function How to distinguish 2) and 3)?
Lineage analysis suggest: Many cells that normally die now survive ced-3 and ced-4 are involved in cell killing How do ced-3 and ced-4 kill the cells? Cells die by murder? Cells die by suicide? cells die by aging? Cells die because of injuries? Cells die by sickness? How to distinguish these possibilities?
Mosaic Analysis Using specific techniques to create genetically mosaic worms, in which some cellscarry a wild-type gene and others are homozygous mutant. Analysis of such animals canreveal where in the animal a gene is needed for its functions How to generate C. elegans genetic mosaics? Mosaic animals have been generated by the spontaneousmitotic loss of an extrachromosomal genetic element that carriesthe wild-type allele of a gene in an otherwise homozygous mutantbackground. free chromosome fragments extrachromosomalarrays
Cells Die by Suicide Rather Than Murder • Yuan and Horvitz demonstrated by mosaic analysis that ced-3 and ced-4 function in the dying cells to kill. • ced-3 encodes a protein with homology with IL-1b converting enzyme (ICE), a cysteine protease. • ced-4 encodes a protein similar to apoptotic protease-activating factor (Apaf-1).
cps-6 ceh-30 CEM cells psr-1 wah-1
Caspases Are Cell Death Executors • Yuan’s group using cell culture experiments showed ICE and CED-3 can both kill in mammalian cells • CED-3/ICE define a family of cysteine proteases, named caspases (aspartate-specific proteases),which so far has 16 family members • A caspase is first synthesized as an inactive protease precursor and later activated by specific proteolysis at specific aspartate residues.
Caspase Family caspase Adapted from Thornberryand Lazebnik, Science 281, 1312-1216, 1998
Stucture of Caspase-3 (CPP32) Adapted from Thornberryand Lazebnik, Science 281, 1312-1216, 1998
Activation of Caspases 1) By self-activation 2) By another cysteine protease or caspase
Discovery of Bcl-2 • In 1986, three groups independently cloned bcl-2 oncogene. bcl-2 oncogene causes follicular lymphoma and is a result of chromosome translocation [t(14;18] that has coupled the immunoglobulin heavy chain locus to a chromosome 18 gene denoted bcl-2 • In 1988, Vaux, Cory, and Adams discovered bcl-2 oncogene causes cancer by inhibiting lymphocyte cell deaths, providing the first evidence that cancer can result from inhibition of cell death • 1990, Stanley Korsmyer’s group showed Bcl-2 localized to mitochondria
C. elegansced-9 Gene Is a Functional Homologue of Bcl-2 • A gain-of-function mutation in ced-9 protects against all cell deaths in nematodes, while loss-of-function mutations cause massive ectopic cell deaths • ced-9 encodes a protein similar to Bcl-2 • Bcl-2 inhibits cell death in nematodes and can partially substitute for ced-9 • CED-9 is localized at mitochondria
Bcl-2/ced-9 Define a Family of Cell Death Regulators • Korsmyer’s group purified a protein, Bax, that associates with and modulates the activity of Bcl-2. Bax by itself can also cause apoptosis in a Bcl-2-independent and caspase-independent pathways. • Thompson’s group identified a gene, named bcl-x, which can be alternatively spliced to generate two proteins that have opposite functions in apoptosis. The long form (Bcl-xL) inhibits apoptosis and the short form (Bcl-xs) cause cell death. • Korsmyer’s group identified another Bcl-2-interacting and death inducing-protein, Bid, which only has one Bcl-2 homology domain (BH3). • Subsequently, more Bid-like death-inducing proteins were identified, all of which has only one BH3 domain. This protein family was called BH3-only Bcl-2 subfamily.
Anti-apoptotic Bcl-2 family members Adapted from Adams and Cory, Science 281, 1322-1226, 1998
Pro-apoptotic Bcl-2 family protein Adapted from Adams and Cory, Science 281, 1322-1226, 1998
Key Features of Bcl-2 Family Proteins • They localize either inducibly or constitutively to outer membranes of mitochondria and nuclei, and membranes of ER • They are capable of forming heterodimers with other family members, especially those with an amphipathic helical BH3 domain • They are capable of forming ion-conducting channels on synthetic membranes
Structures of Bcl-xL and Bid BH3 BH3 Adapted from Chou et al., Cell, 1322-1226, 1999
Puzzles About Bcl-2 • Do they need to localize to mitochondria to function? • Do they need to dimerize with other family members or apoptotic regulators to function? • Do they function as channels to regulate cell death?
Bcl-2 Family Proteins and Cancer • Overexpression of Bcl-2 caused follicular lymphomas • Mutations in Bax cause human gastrointestinal cancer and some leukemias. • In many tumor cell lines, the expression levels of pro- and anti-apoptotic Bcl-2 family members are altered.