Download

1 / 47
570 likes | 951 Vues
● 遗传密码 —— 三联子 ● tRNA 的结构、功能与种类 ● 核糖体的结构与功能 ● 蛋白质合成的过程 ● 蛋白质的运转机制. Contents. (一) 核糖体的结构. 三、核糖体的结构与功能. 1/3的蛋白和2/3的RNA(原核) 2/5的蛋白和3/5的RNA (真核). 核糖体的组成. 36. 表 14-7 原核和真核生物核糖体的组成及功能 核糖体亚基 rRNAs 蛋白 RNA 的特异顺序和功能 细菌

E N D
● 遗传密码——三联子 ● tRNA的结构、功能与种类 ● 核糖体的结构与功能 ● 蛋白质合成的过程 ● 蛋白质的运转机制 Contents
(一)核糖体的结构 三、核糖体的结构与功能 1/3的蛋白和2/3的RNA(原核) 2/5的蛋白和3/5的RNA(真核)
核糖体的组成 36
表14-7 原核和真核生物核糖体的组成及功能 核糖体亚基 rRNAs 蛋白 RNA的特异顺序和功能 细菌 70S 50S 23S=2904b 36种(L1-L36) 2.5×106D 5S=120b 含CGAAC和GTψCG互补 66%RNA 30S 16S=1542b 21种(S1-S21) 16S RNA(CCUCCU)和S-D 顺序(AGGAGG)互补 哺乳动物 80S 60S 28S=4718b 49种 有GAUC和tRNAfMat的TψCG互补 4.2×106D 5S=120b 60%RNA 5.8S=160b 40S 18S=1874b 33种 和Capm7G结合
39 原核生物rRNA 5SrRNA:有两个高度保守的区域 CGAAC与tRNA上的TΨC臂上的GTΨCG识别 GCGCCGAAUGGUAGU与23SrRNA互补,是与 50S核糖体大亚基作用的位点 16SrRNA 3’端ACUCCUUA与mRNA5‘端富含嘌呤的序列 (SD序列)互补 靠近3’端还有一段与23SrRNA互补的序列,在 30S与50S亚基的结合中起作用
40 原核生物rRNA 23SrRNA 存在与tRNAMet序列互补的片段,表明大亚基可能与 tRNAMet的结合相关 近5’端有与5SrRNA互补的片段,表明5S和23S rRNA 存在相互作用
41 真核生物rRNA 5.8SrRNA CGAAC与tRNA上的TΨC臂上的GTΨCG识别 与5SrRNA功能相似 18SrRNA 酵母的18SrRNA的3’端与大肠杆菌16S rRNA具有广泛的同源性 28SrRNA 功能还不清楚
43 三个tRNA结合位点 A位点 Aminoacylsite 氨酰的位点 P位点 Peptidylsite 成肽的位点 E位点 Exitsite 退出的位点
在单个核糖体上,可化分多个功能活性中心, 在蛋白质合成过程中各有专一的识别作用和功能。 ●mRNA结合部位——小亚基 ●结合或接受AA-tRNA部位(A位)——大亚基 ●结合或接受肽基tRNA的部位(P位)——大亚基 ●肽基转移部位——大亚基 ●形成肽键的部位(转肽酶中心)——大亚基 ●多肽链转移到AA-tRNA上释放tRNA的位点(E位)
45 核糖体的功能 蛋白质合成的工厂 包含多个活性中心:结合mRNA的,结合 tRNA的三个位点,结合各种延伸因子的 对模板mRNA进行序列特异性识别
● 遗传密码——三联子 ● tRNA的结构、功能与种类 ● 核糖体的结构与功能 ● 蛋白质合成的过程 ● 蛋白质的运转机制 Contents
氨基酸的活化 翻译的起始 肽链的延伸 肽链的终止 蛋白质前体的加工 四、蛋白质合成的过程
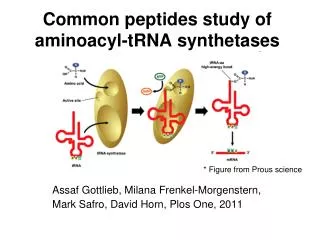
氨基酰-tRNA合成酶 氨基酸 + tRNA 氨基酰- tRNA ATP AMP+PPi (一)氨基酸的活化 氨基酸与对应的tRNA结合 tRNA的3’末端-OH与氨基酸的COOH发生脱 水反应,生成酯结构 ---CCA-OH+HOOC-CH(R)NH2→ ---CCA-OOC-CH(R)NH2
第一步反应 氨基酸活化 AA+ATP+酶(E)→E-AA-AMP+PPi
第二步反应 氨酰基转移 E-AA-AMP+tRNA→AA-tRNA+E+AMP
tRNA与酶结合的模型 tRNA ATP 氨基酰-tRNA合成酶
氨基酰-tRNA合成酶对底物氨基酸和tRNA都有高度特异性。氨基酰-tRNA合成酶对底物氨基酸和tRNA都有高度特异性。 同一氨酰合成酶可以把相同氨基酸加到两个或者更多的带有不同反密码子的tRNA上 氨基酰-tRNA的表示方法: Ala-tRNAAla Ser-tRNASer Met-tRNAMet
原核生物中,起始氨基酸是: 起始AA-tRNA是: 真核生物中,起始氨基酸是: 起始AA-tRNA是: 甲酰甲硫氨酸 fMet-tRNAfMet 甲硫氨酸 Met-tRNAiMet 注意区分两种Met-tRNA Met-tRNAiMet Met-tRNAMet 起始氨酰tRNA 普通氨酰tRNA
原核生物(细菌)为例: 所需成分: 30S小亚基、 50S大亚基、模板mRNA、 fMet-tRNAfMet、GTP、Mg2+ 翻译起始因子:IF-1、IF-2、IF-3、 (二)翻译的起始
翻译起始(翻译起始复合物形成) 可被分成3步: 1. 30S小亚基首先与IF-1, IF-3结合,然后核糖体大小亚基分离 IF-1 IF-3
5' 3' A U G 30S小亚基通过SD序列与mRNA模板相结合。 IF-1 IF-3
SD序列 mRNA中用于结合原核生物核糖体的序列。SD序列在细菌mRNA起始密码子AUG上游10个碱基左右处,有一段富含嘌呤的碱基序列,能与细菌16S rRNA 3’端富含嘧啶区识别,帮助从起始AUG处开始翻译。 5’-AGGAGGU-3’
IF-2 GTP 5' 3' A U G 2. 在IF-2和GTP的帮助下, fMet-tRNAfMet进入小亚基的P位,tRNA上的反密码子与mRNA上的起始密码子配对。 IF-1 IF-3
5' 3' A U G 3、带有tRNA、mRNA和3个翻译起始因子的小亚基复合物与50S大亚基结合,GTP水解,释放翻译起始因子。 IF-2 GTP Pi GDP IF-1 IF-3
IF-2 GTP 5' 3' A U G 原核生物翻译的起始过程 IF-2 Pi -GTP GDP IF-3 IF-1
真核生物翻译起始的特点 ● 核糖体较大, 为80S ● 起始因子比较多 ● mRNA 5′端具有m7Gppp帽子结构 ● Met-tRNAMet ● mRNA的5′端帽子结构和3′端polyA都参与形成翻译起始复合物
原核生物中30S小亚基首先与mRNA模板相结合,再与fMet-tRNAfMet结合,最后与50S大亚基结合。而在真核生物中,40S小亚基首先与Met-tRNAMet相结合,再与模板mRNA结合,最后与60S大亚基结合生成80S·mRNA·Met-tRNAMet起始复合物。 真核生物翻译起始复合物形成(区别原核生物)
40S elF-3 40S mRNA ② ATP met elF4E, elF4G, elF4A, elF4B,PAB ③ Met Met-tRNAMet-elF-2-GTP ADP+Pi 60S Met elF-5 各种elF释放 eIF-2B、eIF-3、eIF-6 ④ ① GDP+Pi 60S Met 真核生物翻译起始复合物形成过程
12 40S小亚基识别起始密码子的“扫描模型” • 40 S小亚基先结合在mRNA 5‘端的任何序列上 • 沿mRNA移动 • 直至遇到AUG发生稳定的相互作用,最后与60 S亚基结合
肽链延伸由许多循环组成,每加一个氨基酸就是一个循环,每个循环包括:AA-tRNA与核糖体结合、肽键的生成和移位。 延伸因子(elongation factor, EF) : 原核生物:EF-T (EF-Tu, EF-Ts) EF-G 真核生物:EF-1 、EF-2 (三)肽链的延伸
1、AA-tRNA与核糖体A位点的结合 需要消耗GTP,并需EF-Tu、EF-Ts两种延伸因子
第二个AA-tRNA在延伸因子 EF-Tu及GTP的作用下,生成 AA-tRNA/EF-Tu/GTP复合物, 然后结合到核糖体的A位。 (只有fMet-tRNAfMet能与 第一个P位点结合,其他的tRNA 必须通过A位点到P位点)
GTP水解释放,通过延伸因子EF-Ts再生GTP, 形成EF-Tu·GTP复合物 EF-Tu-GTP+ EF-Ts EF-Tu-Ts + GDP + Pi EF-Tu-Ts + GTP EF-Tu-GTP + EF-Ts 重新参与下一轮循环
2、肽键形成是由转肽酶/肽基转移酶催化A位上的AA-tRNA转移到P位,与P位的fMet-tRNAfMet上的氨基酸形成肽键,起始tRNA在完成使命后离开核糖体P位点2、肽键形成是由转肽酶/肽基转移酶催化A位上的AA-tRNA转移到P位,与P位的fMet-tRNAfMet上的氨基酸形成肽键,起始tRNA在完成使命后离开核糖体P位点
核糖体向mRNA3’端方向移动一个密码子。 需要消耗GTP,并需EF-G延伸因子 3、移位
二肽酰-tRNA2从A位进入P位,去氨酰-tRNA 被挤入E位, mRNA上的第三位密码子则对应于A位。 延长因子EF-G有转位酶( translocase )活性,可结合并水解1分子GTP,促进核蛋白体向mRNA的3'侧移动 。
Tu GTP 5' 3' A U G fMet fMet
(四)肽链的终止 原核肽链合成终止过程 出现终止密码子 在核糖体的A位点时 释放因子结合密码子 水解P位上多肽链与tRNA 间的二酯键,多肽链释放, 大小亚基解离,蛋白质 合成结束
22 释放因子 释放因子 I类能识别终止密码子,催化多肽链的解放 II类刺激I类从核糖体解离 原核生物的释放因子 I类RFI识别UAG和UAA;RF2识别UGA和UAA II类RF3与核糖体的解离有关,能刺激RF1和RF2 活性 真核生物的释放因子 I类:eRF1能识别UAGUAA和UGA II类:eRF3
25 (五)蛋白质前体的加工 N端fMet或Met的切除:脱甲酰化酶 形成二硫键 特定氨基酸的修饰 磷酸化–OH的反应P137图4-21a 糖基化膜蛋白和分泌蛋白 甲基化细胞质机制内 乙酰化,泛素化,羟基化和羧基化等 切除非功能片段
1、N端fMet或Met的切除 新生蛋白质经蛋白酶切后变成有功能的成熟蛋白质
2、二硫键的形成 两个半胱氨酸-SH-SH-SH二硫键 氧化
3、特定氨基酸的修饰 磷酸化、糖基化、甲基化、乙基化、羟基化和羧基化
4、切除新生肽链中非功能片段 前胰岛素原蛋白翻译后成熟过程示意图
28 (六)蛋白质的折叠 所有新生肽链必须通过正确折叠才能形成动力 学和热力学稳定的三维构象,从而表现出生物 学活性或功能 分子伴侣(molecular chaperone):辅助多肽链正确 折叠、组装、运转和降解的蛋白质,防止或消除肽链的 错误折叠,本身不参与最终产物的形成。 热休克蛋白促进自发折叠的蛋白正确折叠 伴侣素为非自发折叠的蛋白提供正确折叠的微环 境
(七)蛋白质合成抑制剂 青霉素、四环素和红霉素只和原核细胞核糖体作用,广泛应用于人类医学 氯霉素和嘌呤霉素既能与原核细胞核糖体结合,又能与真核细胞核糖体结合, 限制应用