Download

1 / 32
330 likes | 1.26k Vues
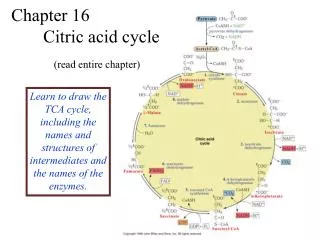
The Tricarboxylic Acid Cycle Krebs’ Cycle Citric Acid Cycle. Hans Krebs. Citric acid cycle overview. The principal gateway to aerobic metabolism . Important source of building blocks for amino acids, glucose, nucleotide bases, cholesterol and porphyrin (organic part of heme).

E N D
The Tricarboxylic Acid Cycle Krebs’ CycleCitric Acid Cycle Hans Krebs
Citric acid cycle overview • The principal gateway to aerobicmetabolism. • Important source of building blocks for amino acids, glucose, nucleotide bases, cholesterol and porphyrin (organic part of heme). • It is the condensation of a 4-carbon molecule (oxaloacetate) with a 2-carbon acetyl unit (acetyl CoA) to yield a 6-carbon tricarboxylic acid (citrate), which is sequentially decarboxylated and oxidised to regenerate oxaloacetate
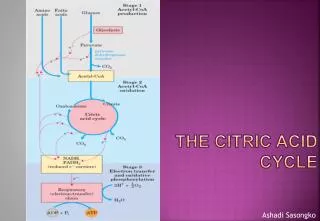
Citric acid cycle overview • Its overall function is to harvest high energyelectrons from carbon fuels. • Net yield/cycle: 2 CO2 + 3 NADH + 1 FADH2 • (note: no ATP made or O2 needed). • The 8 e- carried by NADH and FADH2 are destined for oxidative phosphorylation. • Oxidative phosphorylation90-95% of ATP formed.
Functions of TCA cycle • Energy provision • Energy from carbohydrate, fat & protein • Final oxidation of Carbon compounds to CO2 & H2O • Reduced cofactors (NADH & FADH2) for oxidative phosphorylation • Substrate for biosynthesis of macromolecules • Substrate for glucose synthesis • Substrate for amino acid/protein • Substrate for porphyrin & haem (haem comprises porphyrin and iron)
Glucose Cytoplasm Glycolysis ATP Pyruvate CO2 Pyruvate Mitochondrion PDH acts here Acetyl CoA NADH Krebs’ cycle Electron transport system GTP ATP How many ATPs from 1 molecule of glucose?
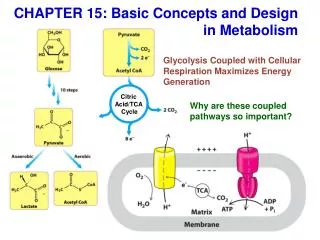
Link between glycolysis and citric acid cycle • Glycolysis ends with formation of pyruvate in the cytoplasm • Pyruvate transported to the mitochondrial matrix where it is oxidised and decarboxylated by the pyruvate dehydrogenase complex (PDH) to form acetyl CoA. • Net rxn: Pyruvate + NAD+ + CoA acetyl CoA + CO2 and NADH • Conversion of pyruvate to acetyl CoA is an irreversible rxn, with capture of high transfer potential electrons in NADH.
Substrates for TCA • Acetyl Coenzyme A • From pyruvate & glycolysis • From fatty acid breakdown, b-oxidation • From ketone bodies • From ketogenic amino acids • Other carbon sources • a-ketoglutarate from amino acid breakdown • Succinyl CoA from propionate (Volatile Fatty Acid – very important in RUMINANTS) • Oxidised cofactors • NAD+ and FAD (TCA operates under aerobic conditions because these electron acceptors are regenerated when NADH& FADH2 t’fer e- to O2 in e-transport chain) • GDP and pi
Products of TCA cycle • 2 x CO2 released in breath • 3 x NADH Ox Phos (3ATP) • 1 x FADH2 to Ox Phos (2ATP) • GTP
Location of TCA cycle • Tissue location • All tissues with mitochondria • Not in Red Blood Cell • Intracellular location • In mitochondria • In matrix of mitochondria • Mitochondrial origin • Synergistic bacteria • Has its own DNA
Outer membrane Inner membrane TCA Cristae Matrix Mitochondria & TCA cycle • Outer membrane • porous • Inner membrane • Impermeable: controlled entry • Inter-membrane space • Matrix • Location of TCA cycle • Cristae
Physiology of the TCA Cycle • Up-regulation • Increased demand for energy • Exercise • Aerobic when oxygen supply is balanced to demand • Anaerobic when oxygen supply can not meet demand • Temperature decrease in environment • Disease • Down-regulation • Rest, hibernation, recovery
Control of TCA Cycle • Whole body control • Endocrine & CNS control, exercise increases • Environmental effectors, fight or flight • Cellular control • Energy levels • ATP/ADP(AMP) ratios • NADH/NAD+ ratios • Regulatory enzymes responding to metabolites
Features of the TCA cycle • 4 carbon acid (oxaloacetate) combines with 2 carbon unit acetyl group of acetyl CoA • Over further 7 reactions 2 x CO2 released • This is a net oxidation of the carbon as in burning wood • Released energy is kept under control, converted to useful energy (ATP) not heat & light • Oxaloacetate is restored at end of the 7 reactions!
BC: Before Cycle Irreversible funneling of pyruvate (product of glycolysis) into TCA cycle • Pyruvate can cross inner mitochondria membrane • Converted by Pyruvate Dehydrogenase, a multi enzyme complex that requires 5 co-enzymes*: • thiamine pyrophosphate (from thiamine [vit B1]), • lipoic acid (another vitamin of vitamin B complex), • Co A (from pantothenic acid), • FAD (from riboflavin) • NAD+ (from nicotinamide) • 3 reaction process • CO2 released • NAD+ reduced to NADH and H +
coenzyme A is carrier of acetyl groups + Decarboxylation Pyruvate Dehydrogenase: a multi-enzyme complex Transfer of acetyl group Transfer of acetyl group
Steps in citric acid cycle • The first step is the condensation of oxaloacetate (C4) and acetyl CoA (C2) to 6-carbon citrate, catalysed by citrate synthase. • Next, citrate is isomerised to isocitrate by aconitase, an iron-sulphur enzyme.
Steps in citric acid cycle • In turn, isocitrate is oxidised and decarboxylated to -ketoglutarateby isocitrate dehydrogenase. The 1st pair of high energy electrons formed (NADH). • Isocitrate dehydrogenase inhibited by: • ATP and NADH when cell is energy rich • Leads to: • Excess isocitrate • Excess citrate (substrate for Fatty acid synthesis)
Steps in citric acid cycle • -Ketoglutarate is also oxidised and decarboxylated to succinyl CoA, mediated by the -ketoglutarate dehydrogenase complex (homologous to the PDH complex). Succinyl CoA formed with a 2nd NADH.
Steps in citric acid cycle • Cleavage of thioester bond of succinyl CoA is coupled to phosphorylation of GDP to form GTP and succinate, catalysed by succinyl CoA synthase. • Energy release from thioester bond used to phosphorylate GDP. Substrate level phosphorylation
Steps in citric acid cycle • Finally, succinate is regenerated to oxaloacetate in 3 steps: oxidation (succinate dehydrogenase), hydration (fumarase) and a second oxidation (malate dehydrogenase). Energy yield from these 3 steps: FADH2 and a (3rd) NADH.
Succinate dehydrogenase • Membrane bound on inner mitochondrial membrane • Domain in matrix to react with succinate • Contains FAD as cofactor (free E change insufficient to reduce NAD+) • Trans double bond formed in fumarate • FADH2enters electron transport chain & Oxidative Phosphorylation
Glucose Cytoplasm Glycolysis ATP Pyruvate CO2 Pyruvate Mitochondrion PDH acts here Acetyl CoA NADH Krebs’ cycle Electron transport system GTP ATP Control of pyruvate dehydrogenase complex
Glucose Pyruvate Acetyl CoA CO2 Lipids Control of pyruvate dehydrogenase complex • In animals, form’n of Acetyl Co A from pyruvate is key irreversible step because they are unable to convert Ac CoA into glucose. • Oxidative decaryboxylation of pyruvate to acetyl CoA commits carbon atoms of glucose to 2 fates: • Oxidation to CO2 by TCA cycle (& generation of E) OR Incorporation into lipid • PDH complex at critical decision point in metabolism • TIGHTLY REGULATED
Control of pyruvate dehydrogenase complex • Inhibited by products of PDH i.e. high levels of Acetyl CoA & NADH. CoA and NAD+ reverse the inhibitory effects • Controlled by E charge. PDH component (of the complex) is inhibited by GTP and activated by ADP. • Regulation by reversible phosphorylation. • Inactive when serines are phosphorylated (by PDH kinase family members) • Active when serines are dephosphorylated (by PDH phosphatase family members). • Insulin stimulates dephosphorylation (accelerates pyruvate to Ac CoA and hence glucose to pyruvate)
GTP • Rate adjusted to meet cell’s need for ATP • ATP is allosteric inhibitor of citrate synthase • ADP allosterically stimulates isocitrate dehydrogenase • High E charge inhibits a-ketoglutarate dehydrogenase
Rate of TCA cycle adjusted to meet cell’s need for ATP • Activity of citrate synthase • ATP is allosteric inhibitor – increases KM for acetyl CoA • Activity of isocitrate dehydrogenase: • Inhibited by NADH (displaces NAD+)and ATP. • Activated by NAD+ (enhances affinity for substrates) and ADP. • Activity of -ketoglutarate dehydrogenase: • Inhibited by succinyl CoA, NADH (products of the reaction it catalyses) and ATP (high energy charge).
Energy yield comparison • Net stoichiometry from electron transport chain (oxidative phosphorylation) is about 3 ATP per NADH, and 2 ATP per FADH2. • 1 round citric acid cycle produces 1 GTP, 3 NADH and 1 FADH2, equivalent to about 12.0 ATP. 1 acetyl unit generates 12 ATP molecules. Also 1 NADH from pyruvate to acetyl CoA conversion. Hence, total of 15 ATP from pyruvate to water. • In glycolysis, 1 glucose molecule generates 2 acetyl CoA, and a net of 2 ATP.
Energy from TCA cycle Before Cycle
Maintaining the TCA cycle • TCA intermediates can be used for biosynthesis • Net loss of substrate reduces rate of cycle & ATP production • Replenished by ‘anaplerotic reactions’ An enzyme-catalysed chemical reaction that recharges the supply of intermediatemolecules • Oxaloacetate is used for gluconeogenesis especially in ruminants and carnivores • Pyruvate is converted to oxaloacetate by pyruvate carboxylase reaction (see gluconeogenesis)
Biosynthetic building blocks from TCA Glucose via gluconegenesis
Oxaloacetate and gluconeogenesis • Gluconeogenesis from pyruvate is not the direct reversal of glycolysis. • In the mitochondria, pyruvate is carboxylated by pyruvate carboxylase (PC) to oxaloacetate. PC is activated in the presence of acetyl CoA. • If oxaloacetate is abundant or if glucose is needed, gluconeogenesis proceeds via malate conversion for transport into the cytoplasm. pyruvate + CO2 + ATP + H20 oxaloacetate + ADP + Pi + 2H+ Pyruvate Carboxylase