Download

1 / 72
720 likes | 736 Vues
Nucleic acids: metabolism II and DNA replication. Andy Howard Biochemistry Lectures, fall 2010 1 December 2010. Nucleic Acids and Replication. We’ll finish biosynthesis and salvage pathways for nucleotides We’ll briefly discuss DNA structure Then we’ll discuss DNA replication.

E N D
Nucleic acids:metabolism II andDNA replication Andy HowardBiochemistry Lectures, fall 20101 December 2010 Nucleotide metab 2+replication
Nucleic Acids and Replication • We’ll finish biosynthesis and salvage pathways for nucleotides • We’ll briefly discuss DNA structure • Then we’ll discuss DNA replication Nucleotide metab 2+replication
Purine synthesis Path to IMP IMP to AMP and GMP Deoxyribonucleotides Ribonucleotide reductase dTMP Salvage pathways Purines Pyrimidines Nucleotide Catabolism DNA structure Prokaryotic replication Semiconservative replication Unwinding of parent DNA Leading-strand replication Lagging-strand replication Okazaki fragments What we’ll cover Nucleotide metab 2+replication
Purine synthesis • Considerably more complex than pyrimidine synthesis • More atoms to condense and two rings to make • More ATP to sacrifice during synthesis • Several synthetase (ligase) reactions require ATP • Based on PRPP, gln, 10-formyl THF, asp Nucleotide metab 2+replication
PRPP + gln to phospho-ribosylamine • PRPP aminated:PRPP + gln glu + PPi +5-phospho--D-ribosylaminevia glutamine-PRPP amidotransferase • transferase structure • Product is unstable(lasts seconds!) 1 PDB 1ECF120 kDa tetramerdimer shownE.coliEC 2.4.2.14, 2Å Nucleotide metab 2+replication
Phospho-ribosylamine to GAR • Amine condenses with glycine to form glycinamide ribonucleotide (GAR) • ATP hydrolysis drives GAR synthetase reaction to the right 2 PDB 2YRX50 kDa monomerGeobacillus kaustophilusEC 6.3.4.13, 1.9Å Nucleotide metab 2+replication
FGAR Formylation of GAR 3 • 10-formyl THF donates a formyl (-CH=O) group to end nitrogen with the help of GAR transformylase to form formylglycinamide ribonucleotide (FGAR) • Rossmann PDB 1MEO47 kDa dimerEC 2.1.2.21.72Åhuman Nucleotide metab 2+replication
FGAM FGAR to FGAM • Glutamine sidechain is source of N for C=O exchanging to C=NH via FGAM synthetase to form formylglycinamidine ribonucleotide (FGAM):FGAR + gln + ATP + H2O FGAM + glu + ADP + Pi 4 PurS component ofFGAM synthetasePDB 1GTD40 kDa tetramerMethanobacterium2.56Å; EC 6.3.5.3 Nucleotide metab 2+replication
Aminoimidazoleribonucleotide FGAM to AIR • Cyclize FGAM to aminoimidazole ribonucleotide • ATP drives the AIR synthetase reaction: • FGAM + ATP + H2O AIR + ADP + Pi • E.C. in Wikipedia was wrong until this morning:it should be 6.3.3.1 5 • PDB 2V9Y147 kDa tetramerdimer shownhuman, 2.1Å Nucleotide metab 2+replication
CAIR AIR to CAIR 6 • AIR is carboxylated;expenditure of an ATP:AIR + HCO3- + ATP carboxyaminoimidazole ribonucleotide + ADP + Pi + 2H+ • AIR carboxylase • E.coli version is two enzymeseukaryotes havea single enzyme • No cofactors! PDB 2NSH149 kDa octamermonomer shownEC 4.1.1.21, 2.1Å E.coli Nucleotide metab 2+replication
CAIR+asp to SAICAR • CAIR + asp + ATP aminoimidazole succinylocarboxamide ribonucleotide + ADP + Pi • Enzyme is SAICAR synthetase • Domain 1: homolog ofphosphorylasekinase • Domain 2: ATP-binding 7 PDB 2CNQ34 kDamonomeryeast, 1.8ÅEC 6.3.2.6 Nucleotide metab 2+replication
SAICAR to AICAR • SAICAR aminoimidazole carboxamide ribonucleotide + fumarate • Enzyme is adenylosuccinate lyase • Net result of two reactions is just replacing acid with amide; • That’s like first 2 reactions in urea cycle, except ADP, not AMP, is the product 8 PDB 2PTR, EC 4.3.2.21.85Å, 203 kDa tetramerdimer shown; E.coli Nucleotide metab 2+replication
AICAR to FAICAR • 10-formylTHF donates HC=O:AICAR + 10-formylTHF formamidoimidazole carboxamide ribonucleotide + THF • Enzyme: AICAR transformylase • Like step 3 • Generally a bifunctional enzyme combined with next step • This part is like cytidine deaminase (see below) 9 PDB 1THZ, 1.8Å130 kDa dimer, chickenEC 2.1.2.3+3.5.4.10 Nucleotide metab 2+replication
FAICAR to IMP • We made it:FAICAR inosine 5’-monosphosphate + H2O • Bifunctional enzyme; this part is called IMP cyclohydrolase or inosicase • Hydrolase part is like methylglyoxal synthase 10 PDB 1PL0260 kDa tetramerdimer shown; human Nucleotide metab 2+replication
So now we have a purine. What next? • Enzymatic conversions to AMP or GMP;Details on next few slides • AMP and GMP can be further phosphorylated to make ADP, GDP with specific kinases (adenylate kinase and guanylate kinase) • GTP made with broad-spectrum nucleoside diphosphate kinase Nucleotide metab 2+replication
IMP to adenylosuccinate • IMP + aspartate + GTP adenylosuccinate + GDP + Pi • Enzyme is adenylosuccinate synthase • Similar to step 7 in IMP synthesis PDB 2V40, 1.9Å101 kDa dimermonomer shownhumanEC 6.3.4.4 Nucleotide metab 2+replication
Adenylosuccinate to AMP • Adenylosuccinate AMP + fumarate • Like reaction 8 in the IMP pathway; in fact, it uses the same enzyme, adenylosuccinate lyase PDB 2PTR203 kDa tetramerdimer shown; E.coli Nucleotide metab 2+replication
IMP to XMP • IMP + H2O + NAD+Xanthosine monophosphate + NADH + H+ • Enzyme: IMP dehydrogenase • TIM-barrel, aldolase-like protein PDB 1ME8221 kDa tetramer;monomer shownTritrichomonas foetus Nucleotide metab 2+replication
XMP to GMP • XMP + gln + H2O + ATP GMP + glu + AMP + PPi • Enzyme: GMP synthetase • Typical 3-layer sandwich PDB 2DPL68 kDa dimerPyrococcus horikoshii Nucleotide metab 2+replication
Adenylate kinase • Reminder:ATP + AMP 2 ADP • Metal ions play a role in enzyme structure • Enzymes like this need to shield their active sites from water to avoid pointless hydrolysis of ATP PDB 1ZIN24 kDa monomerBacillus stearothermophilus EC 2.7.4.3 1.6Å Nucleotide metab 2+replication
Guanylate kinase • GMP + ATP GDP + ADP • “P-loop”-containing ATP-binding proteins • Rossmann fold • Often membrane-associated and intimately associated with other functions PDB 2QOR22 kDa monomer EC 2.7.4.8Plasmodium vivax 1.8Å Nucleotide metab 2+replication
Purine control I: IMP level • Note that GTP is a cosubstrate in making AMP from IMP • ATP is a cosubstrate in making GMP from IMP • So this helps balance the 2 products Nucleotide metab 2+replication
Purine control II:feedback inhibition • PRPP synthetase inhibited by purines, but only at unrealistic concentrations of [Pur] • Step 1 (gln-PRPP amidotransferase) is allosterically inhibited by IMP, AMP, GMP • Adenylosuccinate synthetase is inhibited by AMP • XMP and GMP inhibit IMP dehydrogenase Nucleotide metab 2+replication
Making deoxyribonucleotides • Conversions of nucleotides to deoxynucleotides occurs at the diphosphate level • Reichard showed that most organisms have a single ribonucleotide reductase that converts ADP, GDP, CDP, UDP to dADP, dGDP, dCDP, and dUDP • NADPH is the reducing agent Nucleotide metab 2+replication
RNR1PDB 1R1R258 kDa dimerE.coli Ribonucleotide reductase heterotetramer • 2 RNR1 subunits; each has • a helical 220-aa domain • 10-strand 480-aa structure (thiols here) • 5-strand 70-aa structure • 2 RNR2 subunits; each has • A diferric ion center • A stable tyrosyl free radical RNR2PDB 1PJ082 kDa dimerE.coli Nucleotide metab 2+replication
Mechanism of RNR (box 18.3) • Y122 in RNR2 is converted to stable free radical • Radical transmitted to RNR1 cys439 • Cys439 reacts with substrate 3’-OH to form free radical at C3’ • Substrate dehydrates to carbonyl at C3’ and free radical at C2’; S- formed at Cys462 • Disulfide formed between Cys462,Cys225;radical regenerated at Cys439 Nucleotide metab 2+replication
Ribonucleotide reductase: control • ATP, dATP, dTTP, and dGTP act as allosteric modulators by binding to two regulatory sites on the enzyme • Activity site (A) regulates activity of catalytic site • When ATP binds at A, activity goes up • When dATP binds at A, activity inhibited overall • Specificity site (S) controls which substrates can be turned over • ATP at A + ATP or dATP at S : pyrimidines only • dTTP at S : activates reduction of GDP • dGTP at S : activates reduction of ADP Nucleotide metab 2+replication
dUDP to dUMP(for making dTMP) • dTMP formed at monophosphate level(from dUMP) • dUMP derived three ways: • dUDP + ADP dUMP + ATP • dUDP + ATP dUTP + ADPdUTP + H2O dUMP + PPi • dCMP + H2O dUMP + NH4+ Nucleotide metab 2+replication
5,10-methylene THF Thymidylate synthase reaction (fig.18.15) • dUMP + 5,10-methyleneTHF dTMP + 7,8-dihydrofolate • Unusual THF reaction in that cofactor gets oxidized as well as giving up a carbon • CH2 from 5,10-methylene group • extra H from C6 • So DHF must be reduced back to THF via DHFR and get its methylene back from SHMT dihydrofolate Nucleotide metab 2+replication
Thymidylate synthase • Generally the controlling step in DNA synthesis because [dTTP] < other [deoxynucleoside triphosphates] • Therefore a target for cancer chemotherapy and other therapies that target rapidly-dividing cells • Enzyme is a 2-layer sandwich PDB 2G8O58 kDa dimerE.coli(with dUMP and cofactor analog) Nucleotide metab 2+replication
Thymidylate synthase and drug design • Both folate analogs and dUMP analogs can interfere with (DHFR SHMT dTMP synthase … ) cycle • 5-fluorouracil is specific to thymidylate synthase Nucleotide metab 2+replication
DHFR • Converts DHF to THF:DHF + NADPH + H+THF + NADP+ • SHMT then converts THF to 5,10-methyleneTHF • 3-layer sandwich PDB 1KMV20 kDa monomerhuman folate Nucleotide metab 2+replication
DHFR as a drug target • Human DHFR inhibition: kill cancer cells faster than healthy cells • Eukaryotic DHFR also catalyzes folate DHF • Prokaryotic DHFR doesn’t; • DHF derived by another mechanism in bacteria • So there are antibacterials based on the structural differencs between prokaryotic and eukaryotic DHFR Nucleotide metab 2+replication
Special case:protozoan TSynth/DHFR • Bifunctional enzyme: • Thymidylate synthase • Dihydrofolate reductase • Presumably some entropic advantage • Maybe electrostatics too, allowing the negative charges on DHF to tunnel through;but cf. Atreya et al (2003) J.Biol.Chem.278:28901. DHFR-TSPDB 1J3K104 kDa dimerPlasmodium falciparum Nucleotide metab 2+replication
Recovery pathway to dTMP • Deoxythymidine can be phosphorylated by thymidine kinase:deoxythymidine + ATP dTMP + ADP • Labeled thymidine is convenient for monitoring intracellular synthesis of DNA because thymidine enters cells easily PDB 1E2K73 kDa monomerHerpes simplex virus Nucleotide metab 2+replication
Fates of polynucleotides • Nucleic acids hydrolyzed to mononucleotides via nucleases • Mononucleotides are dephosphorylated via nucleotidases and phosphatases • Resulting nucleosides are deglycosylated via nucleosideases or nucleoside phosphorylases • Resulting bases are sent either into salvage pathways or get degraded and excreted Nucleotide metab 2+replication
Salvage pathways • We can describe them, and we will: but why do they matter so much? • They provide energy savings relative to denovo synthesis (think of all the ATP we used in making IMP!) • Considerable medical significance to interference with these pathways • Intracellular nucleic acid bases are usually recycled; dietary bases are usually broken down and excess nitrogen excreted Nucleotide metab 2+replication
Orotate phosphoribosyl transferase • Principal salvage enzyme for pyrimidines • Orotate + PRPP OMP + PPi • OMP can then reenter UMP synthetic pathway (decarboxylation to UMP, then form UDP and CDP) • Same enzyme can act on other pyrimidines to make nucleotides:Pyr + PRPP PyrMP + PPi PDB 2PS150 kDa dimerYeastEC 2.4.2.101.75Å Nucleotide metab 2+replication
Pyrimidine interconversions (fig. 18.19) • All phosphorylations & dephosphorylations can and do happen • UTP can be aminated to CTP • CDP and UDP can be reduced to dCDP and dUDP • dCMP can deaminate to dUMP • Cytidine can be converted to uridine • dUMP can be methylated to dTMP Nucleotide metab 2+replication
Purine nucleotide salvage • Two phosphoribosyl transferases convert adenine, guanine, and hypoxanthine to AMP, GMP, and IMP • Adenine phosphoribosyl transferase is specific • HGPRT accepts both hypoxanthine and guanine Hypoxanthine-guaninephosphoribosyl transferasePDB 1FSGEC 2.4.2.8, 1.05Å102 kDa tetramerdimer shownToxoplasma gondii Nucleotide metab 2+replication
Purine Interconnections(fig. 18.18) • All phosphorylations and dephosphorylations can and do occur • ADP and GDP can be reduced to dADP and dGDP • AMP can deaminated to IMP (new) • IMP can be aminated to AMP • IMP can oxidized to XMP • XMP can be aminated to GMP • Guanine, adenine can be phosphoribosylated to GMP and AMP Nucleotide metab 2+replication
Fates of CMP and cytidine • CMP’s phosphate can be hydrolyzed off • That’s followed by deamination of cytidine to make uridine • Catalyzed by cytidine deaminase • Another sandwich protein Cytidine deaminasePDB 2FR566 kDa tetramerMouseEC 3.5.4.5, 1.48Å Nucleotide metab 2+replication
Hydrolysis of U, dU and dT • Glycosidic bond in uridine or thymidine is hydrolyzed by phosphate: • Uridine + Pi -D-ribose-1-P + uracil • Enyzme is uridine phosphorylase • Similar enzyme handles deoxyuridine • Similar reaction using thymidine phosphorylase yields thymine + -D-deoxyribose-1-P Uridine phosphorylasePDB 1RXYEC 2.4.2.3, 1.7Å167 kDa hexamer Dimer shown E.coli Nucleotide metab 2+replication
Uracil to acetyl CoA;thymine to succinyl CoA • Reduced to dihydrouracil and dihydrothymine • Hydrated and ring-opened toureidopropionate or ureidoisobutyrate • Eliminate bicarbonate and ammonium to yield -alanine or -aminoisobutyrate • Several reactions from there to acetyl CoA and succinyl CoA Dihydro-pyrimidinasePDB 1GKPEC 3.5.2.21.29Å302 kDa hexamerThermus Nucleotide metab 2+replication
Uric acid Purine catabolism • Nucleoside or deoxynucleoside + phosphate base + (D)-ribose 1-P • Hypoxanthine and guanine both lead to uric acid as a product • Uric acid is the final excreted nitrogenous compound in primates, birds, and some reptiles • Other organisms catabolize it further Nucleotide metab 2+replication
Uric acid Uric acid to allantoin • Urate oxidase:urate + 2H2O + O2 allantoin + H2O2 + CO2 • That’s the final product in a lot of mammals, turtles, some insects, gastropods • Other organisms catabolize allantoin further Allantoin Nucleotide metab 2+replication
Urate oxidase • Catalyzes the decarboxylation of uric acid to allantoin Urate oxidase134 kDa tetramermonomer shownEC 1.7.3.3, 1.5ÅAspergillus flavus Nucleotide metab 2+replication
Lesch-Nyhan syndrome Michael Lesch William Nyhan • Complete lack of hypoxanthine-guanine phosphoribosyl transferase • So hypoxanthine and guanine are degraded to uric acid rather than being built back up into IMP and GMP • Leads to dangerous buildup of uric acid in nervous tissue • Neurological effects are severe and poorly understood Nucleotide metab 2+replication
Sodium urate Gout Sodium urate crystals accumulating • Accumulation of sodium urate and uric acid, both of which are only moderately soluble • Arises from inadequate (~10%) functionality of HGPRT, so that urate accumulates in peripheral tissues, particularly the feet Benjamin Franklin(celebrated gout sufferer) Nucleotide metab 2+replication
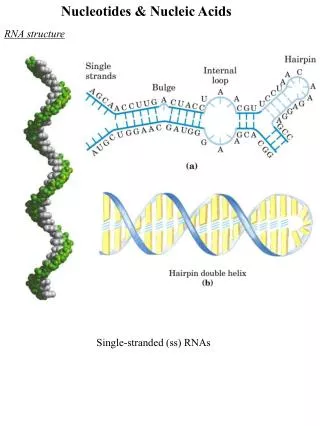
DNA structure • We’ll take a brief sojourn into 3-D structure of DNA • You’ve been told about Watson-Crick base pairing and the double helix since high school • Here we’ll examine the higher levels of structure, including the proteins that enable those higher levels to happen Nucleotide metab 2+replication