Download
1 / 61
840 likes | 1.71k Vues
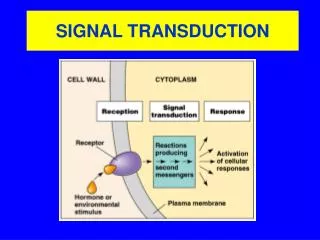
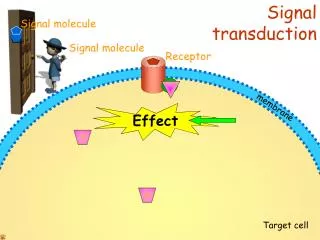
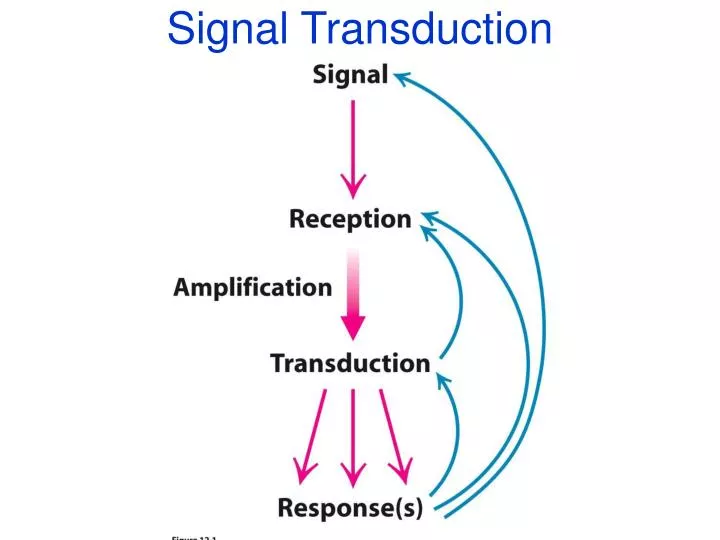
Signal Transduction. Signal Transduction. Signal outside cell is recognized Transmission across membrane Effect inside cell. Signal Transduction & Molecular Circuits. A primary messenger (hormone-H) binds to the extracellular part of a membrane-embedded receptor (R)

E N D
Signal Transduction • Signal outside cell is recognized • Transmission across membrane • Effect inside cell
Signal Transduction & Molecular Circuits A primary messenger (hormone-H) binds to the extracellular part of a membrane-embedded receptor (R) The HR complex generates a second messenger inside the cell which activates proteins that alter the biochemical circuitry inside the cell Then, mechanisms are activated to terminate the signal transduction pathway
Signal Transduction Molecular Switches • Membrane embedded receptor Proteins transmit information into the cell • The Ras protein, like the Ga subunit of heterotrimeric G proteins, cycles between an inactive GDP-bound form and active form bound to GTP • Tyrosine kinase modules of some receptors upon dimerization are activated by cross-phosphorylation. Phosphorylated tyrosines serve as docking sites for adaptor and signaling proteins which permit further propagation of the signal. • Net result is amplification, fidelity and diversity
G-proteins-also known as GPCRsfor G-protein coupled receptors • Trimeric molecular amplifiers (,, subunits, and are lipid-linked thus membrane bound) • Bind to 7-transmembrane receptors on the cytosolic side • Receptor activationswitch GTP for GDP disassociation of from
Switching States • 3 main areas of G-protein change conformation: • Switch I: Moves closer to Guanine when active, Thr177 H-bonds to the -phosphate of GTP • Switch II: a-helix 2 rotates so G199 can H-bond to -phosphate, which pulls b-strand 3 away from b-strand 1 and toward b-strand 2. This breaks old hydrogen bonds and makes new ones. • Switch III: interacts with switch II which propagates its structural changes to it.
Mechanism of Hydrolysis • General idea: • generate OH- to attack -phosphate • Have to neutralize the negative charge on the phosphate
The odd case of Ras • Ras lacks the Arg that stabilizes the negative charge on the phosphate! • Ras is very very slow at hydrolyzing GTP—is this why? • Ras GTPase activating protein (GAP) supplies the necessary Arg residue and speeds up the reaction nearly 100,000 fold
Why is all this important? • Ras is a signalling molecule activated by various receptors in different pathways, including cell growth signals. • Ras is mutated in 25% of human tumors • Mutations inactivate the GTP hydrolysis step, thereby keeping Ras switched “on”—and thus Ras keeps on stimulating cells to divide.
What G-proteins can Do Source: Molecular Biology of the Cell, fourth ed. 2002
The G-protein Cycle • GTP form = active • GDP form = inactive • activated -subunit hydrolyzes GTP to GDP • special proteins help GTP hydrolysis (Regulators of G-protein Signaling, RGS) • and subunits re-assemble into inactive form
Cholera Toxin (CT) • CT is an AB toxin. • B is a pore that allows toxin entry into cells • A is an enzyme that stabilizes the active GTP-bound form of Ga • Active Ga activates Protein Kinase A (PKA) • PKA activates by phosphorylation a Cl- channel and a Na+ - H + exchanger • The net consequence is massive loss of NaCl and water from the intestine
Why use G-proteins? • Amplification at two steps: • Active receptor can activate multiple G-proteins (by causing GTP to replace GDP) • Each downstream target (eg adenylate cyclase) can produce many second messengers (eg, cyclic AMP) • Cells can regulate how long the switch is turned on • Different RGS molecules in different cells or at different times
How Does it Work? • Why does GTP binding cause disassociation of from the subunits? • How does hydrolysis of GTP occur to re-set the system?
The structure of the G subunit • Contains 7 repeats of ~40aa: WD repeats
Phosducin regulates light adaptation in retinal rods • In rod cells: • Rhodopsin absorbs photons • Activates G-protein called transducin • Transducin activates cGMP phosphodiesterase degradation of cGMP • One photondegradation of over 100,000 cGMP molecules!! • To dampen the sensitivity, phosducin reduces transducin’s activity • Binds G and pulls it into the cytoplasm where it is inactive
G phosducin Phosducin blocks G binding site of G Phosducin binding is negatively regulated by phosphorylation of Serine 73
Signal Transduction Molecular Switches • Membrane embedded receptor Proteins transmit information into the cell • The Ras protein, like the Ga subunit of heterotrimeric G proteins, cycles between an inactive GDP-bound form and active form bound to GTP • Tyrosine kinase modules of some receptors upon dimerization are activated by cross-phosphorylation. Phosphorylated tyrosines serve as docking sites for adaptor and signaling proteins which permit further propagation of the signal. • Net result is amplification, fidelity and diversity