Download
1 / 1
10 likes | 76 Vues
Synthesis and Biological Assessment of Sulfonic Acid-Based Glucagon Antagonists Bin Yang, Vasily M. Gelfanov and Richard DiMarchi Department of Chemistry, Indiana University, Bloomington, Indiana 47405-7102, U.S.A. Abstract

E N D
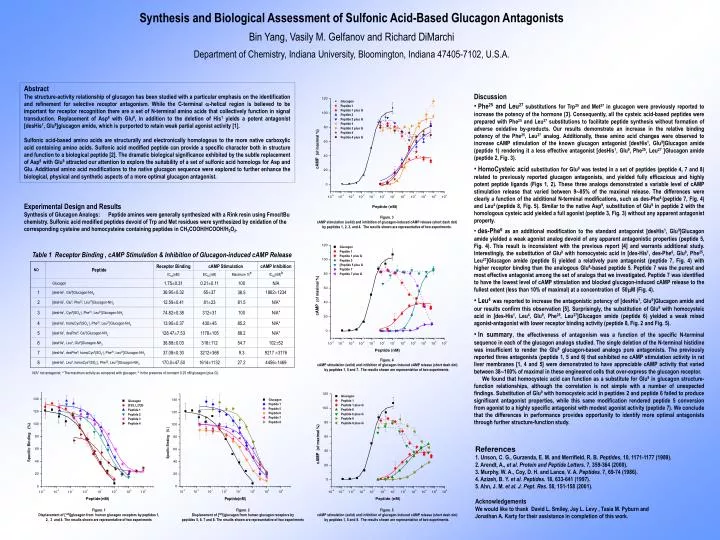
Synthesis and Biological Assessment of Sulfonic Acid-Based Glucagon Antagonists Bin Yang, Vasily M. Gelfanov and Richard DiMarchi Department of Chemistry, Indiana University, Bloomington, Indiana 47405-7102, U.S.A. Abstract The structure-activity relationship of glucagon has been studied with a particular emphasis on the identification and refinement for selective receptor antagonism. While the C-terminal -helical region is believed to be important for receptor recognition there are a set of N-terminal amino acids that collectively function in signal transduction. Replacement of Asp9 with Glu9, in addition to the deletion of His1 yields a potent antagonist [desHis1, Glu9]glucagon amide, which is purported to retain weak partial agonist activity [1]. Sulfonic acid-based amino acids are structurally and electronically homologous to the more native carboxylic acid containing amino acids. Sulfonic acid modified peptide can provide a specific character both in structure and function to a biological peptide [2]. The dramatic biological significance exhibited by the subtle replacement of Asp9 with Glu9 attracted our attention to explore the suitability of a set of sulfonic acid homologs for Asp and Glu. Additional amino acid modifications to the native glucagon sequence were explored to further enhance the biological, physical and synthetic aspects of a more optimal glucagon antagonist. Experimental Design and Results Synthesis of Glucagon Analogs: Peptide amines were generally synthesized with a Rink resin using Fmoc/tBu chemistry. Sulfonic acid modified peptides devoid of Trp and Met residues were synthesized by oxidation of the corresponding cysteine and homocysteine containing peptides in CH3COOH/HCOOH/H2O2. • Discussion • Phe25 and Leu27substitutions for Trp25 and Met27 in glucagon were previously reported to increase the potency of the hormone [3]. Consequently, all the cysteic acid-based peptides were prepared with Phe25 and Leu27 substitutions to facilitate peptide synthesis without formation of adverse oxidative by-products. Our results demonstrate an increase in the relative binding potency of the Phe25, Leu27 analog. Additionally, these amino acid changes were observed to increase cAMP stimulation of the known glucagon antagonist [desHis1, Glu9]Glucagon amide (peptide 1) rendering it a less effective antagonist [desHis1, Glu9, Phe25, Leu27 ]Glucagon amide (peptide 2, Fig. 3). • HomoCysteic acidsubstitution for Glu9 was tested in a set of peptides (peptide 4, 7 and 8) related to previously reported glucagon antagonists, and yielded fully efficacious and highly potent peptide ligands (Figs 1, 2). These three analogs demonstrated a variable level of cAMP stimulation release that varied between 985% of the maximal release. The differences were clearly a function of the additional N-terminal modifications, such as des-Phe6 (peptide 7, Fig. 4) and Leu4 (peptide 8, Fig. 5). Similar to the native Asp9, substitution of Glu9 in peptide 2 with the homologous cysteic acid yielded a full agonist (peptide 3, Fig. 3) without any apparent antagonist property. • des-Phe6as an additional modification to the standard antagonist [desHis1, Glu9]Glucagon amide yielded a weak agonist analog devoid of any apparent antagonistic properties (peptide 5, Fig. 4). This result is inconsistent with the previous report [4] and warrants additional study. Interestingly, the substitution of Glu9 with homocysteic acid in [des-His1, des-Phe6, Glu9, Phe25, Leu27]Glucagon amide (peptide 5) yielded a relatively pure antagonist (peptide 7, Fig. 4) with higher receptor binding than the analogous Glu9-based peptide 5. Peptide 7 was the purest and most effective antagonist among the set of analogs that we investigated. Peptide 7 was identified to have the lowest level of cAMP stimulation and blocked glucagon-induced cAMP release to the fullest extent (less than 10% of maximal) at a concentration of 50M (Fig. 4). • Leu4was reported to increase the antagonistic potency of [desHis1, Glu9]Glucagon amide and our results confirm this observation [5]. Surprisingly, the substitution of Glu9 with homocysteic acid in [des-His1, Leu4, Glu9, Phe25, Leu27]Glucagon amide (peptide 6) yielded a weak mixed agonist-antagonist with lower receptor binding activity (peptide 8, Fig. 2 and Fig. 5). • In summary,the effectiveness of antagonism was a function of the specific N-terminal sequence in each of the glucagon analogs studied. The single deletion of the N-terminal histidine was insufficient to render the Glu9 glucagon-based analogs pure antagonists. The previously reported three antagonists (peptide 1, 5 and 6) that exhibited no cAMP stimulation activity in rat liver membranes [1, 4 and 5] were demonstrated to have appreciable cAMP activity that varied between 38100% of maximal in these engineered cells that over-express the glucagon receptor. • We found that homocysteic acid can function as a substitute for Glu9 in glucagon structure-function relationships, although the correlation is not simple with a number of unexpected findings. Substitution of Glu9 with homocysteic acid in peptides 2 and peptide 6 failed to produce significant antagonist properties, while this same modification rendered peptide 5 conversion from agonist to a highly specific antagonist with modest agonist activity (peptide 7). We conclude that the differences in performance provides opportunity to identify more optimal antagonists through further structure-function study. Figure. 3 cAMP stimulation (solid) and inhibition of glucagon-induced cAMP release (short dash dot) by peptides 1, 2, 3, and 4. The results shown are representative of two experiments. Table 1 Receptor Binding , cAMP Stimulation & Inhibition of Glucagon-induced cAMP Release Figure. 4 cAMP stimulation (solid) and inhibition of glucagon-induced cAMP release (short dash dot) by peptides 1, 5 and 7. The results shown are representative of two experiments. N/A* not antagonist; a The maximum activity as compared with glucagon; b In the presence of constant 0.25 nM glucagon (plus G). References 1. Unson, C. G., Gurzenda, E. M. and Merrifield, R. B. Peptides, 10, 1171-1177 (1989). 2. Arendt, A., et al.Protein and Peptide Letters. 7, 359-364 (2000). 3. Murphy, W. A., Coy, D. H. and Lance, V. A. Peptides. 7, 69-74 (1986). 4. Azizeh, B. Y. et al.Peptides. 18, 633-641 (1997). 5. Ahn, J. M. et al. J. Pept. Res. 58, 151-158 (2001). Acknowledgements We would like to thank David L. Smiley, Jay L. Levy , Tasia M. Pyburn and Jonathan A. Karty for their assistance in completion of this work. Figure. 1 Displacement of [125I]glucagon from human glucagon receptors by peptides 1, 2, 3 and 4. The results shown are representative of two experiments Figure. 2 Displacement of [125I]glucagon from human glucagon receptors by peptides 5, 6, 7 and 8. The results shown are representative of two experiments Figure. 5 cAMP stimulation (solid) and inhibition of glucagon-induced cAMP release (short dash dot) by peptides 1, 6 and 8. The results shown are representative of two experiments.





















