Download

1 / 33
380 likes | 746 Vues
MEMBRANE LIPIDS, PROTEINS & CARBOHYDRATES. The nature of cell membranes Lipids in membranes Proteins in membranes Lipid-protein interactions Carbohydrate presence & roles. Cell membranes:. In the early part of the 20 th century very little was known about cell membranes either at

E N D
MEMBRANE LIPIDS,PROTEINS & CARBOHYDRATES The nature of cell membranes Lipids in membranes Proteins in membranes Lipid-protein interactions Carbohydrate presence & roles
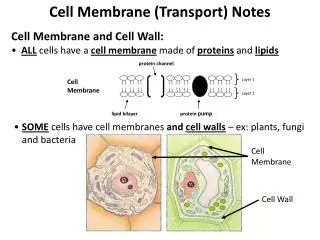
Cell membranes: In the early part of the 20th century very little was known about cell membranes either at the cell surface (plasma membranes) or within the “organs” of cells (sub-cellular organelles). Individuals who pioneered early work in this area were: Overton, Langmuir, Gorter, Grendel, Danielli and Davson.
Overton found (1895) that cell membranes were lipid in nature and not easily penetrated. Langmuir developed a device (1917) to study lipid layers spread out on thin films to examine their air-water interfaces. Gorter & Grendel (1925) first proposed that membranes were made up of lipid bilayers by using Langmuir’s device. Hydrophilic part Hydrophobic part The bilipid model is often incorrectly attributed to Danielli & Davson. The 1925 model (above) failed to account for any proteins associated with the membranes. Danielli & Davson’s contributions will be shown later.
It is now confirmed that the 1925 biliipid layer hypothesis was correct. The lipid portion of the structure is composed of two lipids facing foot-to- foot in which the “feet” (red arrow) are the hydrocarbon tails of the fatty acids and the hydrophilic “heads” (polar head groups) are exposed to an aqueous environment (blue arrows).
BILIPID LAYERS,IN BIOLOGICAL CELLS, ARE FORMED BY THEIR ORIENTATION AGAINST AN AQUEOUS ENVIRONMENT. HOWEVER, BILIPID LAYERS CAN ARRANGE THEMSELVES IN A NONPOLAR ENVIRONMENT AS WELL. BELOW IS ANOTHER ORIENTATION. AIR INSIDE WALL OF SOAP BUBBLE AIR OUTSIDE THE ARRANGEMENT OF THE BILIPID LAYER IS DOMINATED BY THE PREVAILING NEED FOR THE MOST STABLE, LOW ENERGY ASSOCIATION OF THE LIPIDS.
AMPHIPATHIC* LIPIDS SPONTANEOUSLY FORM STRUCTURES IN WATER (BY HYDROPHOBIC BOND FORMATION). THE ARRANGEMENT IS NOTALWAYS A BILIPID LAYER. *LIPIDS WITH A CHARGE AT ONE END OF THE MOLECULE SUCH AS A PHOSPHOLIPID OR A FATTY ACID.
MICELLES..ARE STRUCTURES THAT FORM FROM FATTY ACIDS. THEY ARE SMALL AND HAVE ONLY NON-POLAR LIPID INSIDE. DETERGENTS CAN ALSO FORM MICELLES. BILIPID LAYERS..ARE STRUCTURES THAT FORM FROM PHOSPHOLIPIDS. THEY ARE “LINEAR” AND REMAIN STABLE EXCEPT AT THE EDGES. BECAUSE OF THE “END PROBLEM”, THEY WILL READILY WRAP BACK ON THEMSELVES TO FORM CLOSED VESICLES. LIPOSOMES..ARE SPERICAL VESSELS OF BILIPID LAYERS. THERE MAY BE ONE OR MORE BILIPID LAYERS. CELLS HAVE ESSENTIALLY LIPOSOME COVERINGS WITH A SINGLE BILIPID LAYER. ARTIFICIAL LIPOSOMES MAY BE USED TO DELIVER DRUGS AND REAGENTS FOR DIAGNOSTIC PURPOSES. HOW?
WHAT ABOUT SPHINGOLIPIDS AND CHOLESTEROL IN MEMBRANES? AS PART OF A BILIPID LAYER, SPHINGOLIPIDS FIT IN A MEMBRANE JUST LIKE PHOSOPHOLIPIDS, BUT THEY ALSO CONTRIBUTE DIFFERENT POLAR HEAD GROUPS – ESPECIALLY THE GANGLIOSIDES. THE CARBOHYDRATES ACT AS CELL SURFACE IMMUNOIDENTIFIERS membrane (outer) surface USUAL PART OF THE BILIPID LAYER EXAMPLE: LACTOSYLCERAMIDE
CYTOPLASMIC SIDE CHOLESTEROL IS ACTUALLY A RATHER FLAT MOLECULE AND IS EASILY WEDGED INTO THE ADJOINING PHOSPHOLIPIDS OF A BILIPID LAYER. PHOSPHOLIPID ½ OF BILIPID LAYER
WHEN THE LIPID COMPOSITION IS MODIFIED, THE APPEARANCE & STRUCTURE OF SUCH LAYERS MAY OR MAY NOT BE CHANGED, FOR EXAMPLE, WHEN CHOLESTEROL IS INCORPORATED INTO A BILIPID LAYER NO CHANGE IS OBVIOUS: CHOLESTEROL HOWEVER: THE BILIPID LAYER BECOMES MORE RIGID WITH GREATER CHOLESTEROL INCORPORATION. + CHOLESTEROL
THE LENGTH AND UNSATURATION OF FATTY ACIDS THAT MAKE UP MEMBRANE PHOSPHOLIPIDS MAY HAVE MARKED EFFECTS ON MEMBRANE STRUCTURE AND PROPERTIES. HERE IT IS APPARENT THAT UNSATURATION OF FATTY ACIDS CAUSES TWO EFFECTS: MEMBRANE THINNING AND DISORDER (=INCREASE IN MEMBRANE FLUIDITY). THE EFFECTS PRODUCED ARE ALSO THE SAME WITH AN INCREASE IN MEMBRANE TEMPERATURE. AN INCREASE IN CHAIN LENGTH WOULD HAVE THE OPPOSITE EFFECTS. HOWEVER, ON THE AVERAGE, CHAIN LENGTHS IN MEMBRANES ARE LESS VARIABLE THAN THE DEGREE OF UNSATURATION. ALSO MEMBRANE TEMPERATURES FOR MAMMALS TEND TO BE STABLE. NOTE THAT MEMBRANE FLUIDITY IS AN IMPORTANT PROPERTY IN MEMBRANE FUNCTIONS SUCH AS TRANSPORT.
PROPERTIES OF TRANSITION IN BILIPID MEMBRANES: STATE TRANSITION. This is the property of change in the structure of a membrane. As learned before, phospholipids/ sphingolipids with greater unsaturation form more disorganized bilipids and give a certain “liquidity” to a membrane. Phospho- lipids/sphingolipids at higher temperatures do the same. The conversion of a membrane lipid to a more liquid state ( temp.) occurs at its transition temperature (liquid crystal formation). Liquid crystalline state Paracrystalline or gel state
MOTION TRANSITION Generally, the lipid components of a membrane are under constant motion (vibration). The amount of vibration depends on temperature and composition. DIFFUSION – As a result the lipids tend to spread out and form a uniform distribution on their half of the bilipid layer. Usually they are confined to that layer. However, in more recent studies, flippase enzymes have been described that move phospholipids from one side of a membrane to another. This seem to be required for limited applications such as initial membrane formation or maintenance.
OTHER PROPERTIES THAT LIPIDSIMPOSE ON MEMBRANES ASSYMETRY: THE LIPID COMPOSITION OF THE TRANSVERSE SECTIONS (ACROSS THE BILIPID LAYER) CAN VARY BETWEEN THE OUTSIDE AND INSIDE OF A MEMBRANE. AN EXAMPLE IS SHOWN FOR A RED BLOOD CELL (ERYTHROCYTE). SUCH ASSYMETRY CAN IMPOSE SPECIFIC MEMBRANE CURVATURE OR IMMUNE SPECIFICITY TO A CELL.
ASSYMETRY FROM MEMBRANE TO MEMBRANE IN A CELL: LIPIDS VS. MEMBRANE LOCATION Two extremes in composition of membranes are indicated for mitochondria vs. plasma membranes. In mitochondria, the amount of cholesterol is quite small while that of phosphatidyl ethanolamine is substantial (see arrows). On the other hand, cholesterol is 7x higher in plasma membranes while its phosphatidyl ethanolamine level is only about ½ that of mitochondria. These components contribute to the need for a very fluid membrane in mito- chondria (lack of a stiffening molecule [cholesterol] ) and availability of greater amount of unsaturated fatty acids (found in PE).
THE ROLE OF PROTEINS INMEMBRANES? AT THE TIME THAT DAVSON AND DANIELLI WERE WORKING ON MEMBRANES (1930s – 1940s), THEY REALIZED THAT IT WOULD BE NECESSARY TO EXPLAIN THE ROLE OF PROTEINS IN MEMBRANES TO SHOW HOW – SUBSTANCES WERE TRANSPORTED THROUGH MEMBRANES CELL FUNCTION AND SURVIVAL COULD NOT EXIST WITHOUT SUCH TRANSPORT.
FLUID MOSAIC MODEL ACCOUNTS FOR: 1) 2 KINDS OF PROTEINS- EXTRINSIC & INTRINSIC 2) INCLUSION OF TRANSPORT 3) INTRODUCTION OF CARBOHYDRATES ON PROTEINS
INTRINSIC PROTEINS: ALSO • KNOWN AS INTEGRAL PROTEINS, • ARE FIRMLY ANCHORED IN BILIPID • LAYERS. THESE PROTEINS PASS • THROUGH THE MEMBRANE AND • ARE INVOLVED IN MEMBRANE • FUNCTIONS SUCH AS TRANSPORT • AND HORMONE RECEPTION. • THE EXAMPLE SHOWN IS GLYCO- • PHORIN (A RED BLOOD CELL • PROTEIN) – NOTE: • ALPHA HELIX IN BILIPID LAYER (GREEN ARROW) • 2) CARBOHYDRATES IN OUTER LAYER(RED • ARROW) ABO/ MN BLOOD TYPE MARKER PROTEINS
Na-K-ATPase IS AN INTRINSIC MEMBRANE TRANSPORT ENZYME MECHANISM: (shown at right) It loads up 3 Na+ ions inside the cell ATP causes a decrease in Na+ affinity (d) Na+ release, then K+ uptake occurs (e) dephosphorylation (f) K+ release inside the cell GENERAL DIAGRAM OF Na-K-ATPase
A FEW INTEGRAL (OR INTRINSIC) MEMBRANE PROTEINS EVEN USE b-STRUCTURES (KNOWN AS STRANDS OR BARRELS) TO CROSS CELLULAR MEMBRANES. SOME OTHER EXAMPLES OF INTRINSIC MEMBRANE PROTEINS ARE: SODIUM, POTASSIUM STIMULATED-ATPase (transport of Na and K ions) HUMAN LEUKOCYTE ASSOCIATED (HLA) ANTIGENS (antigen presentation) TolC (outer structure membrane protein of E. coli) [uses b-barrell structure] AQUAPORINS (transport of water molecules) TolC HLA ANTIGEN AQUAPORIN NaK-ATPase
INTEGRAL MEMBRANE PROTEINS PASS THROUGH MEMBRANES WITH ONE OR MORE a-HELICES (sometimes with b-barrels). THOSE PROTEINS THAT ARE INVOLVED WITH TRANSCELLULAR TRANSPORT HAVE BETWEEN 6 AND 12 TRANSCELLULAR HELICES. A HYRODPATHY PLOT MAY BE MADE OF A MEMBRANE PROTEIN’S AMINO ACID SEQUENCE VS. THE HYDROPATHY SCALE OF ITS AMINO ACIDS (e.g. VAL= 4.2 vs. LYS= -3.9 where more positive numbers are more hydrophobic) TO DETERMINE WHERE THE SEQUENCE CROSSES THE CELL MEMBRANE. AS SHOWN, RHODOPSIN WOULD HAVE SEVEN TRANSMEMBRANE a-HELICES.
EXTRINSIC PROTEINS. ALSO CALLED PERIPHERAL PROTEINS. THESE ASSOCIATE WITH THE MEMBRANE BY ELECTROSTATIC INTERACTIONS AND HYDROGEN BONDING. THEY ARE ISOLATED BY MILD TREAT- MENTS SUCH AS SALTING OUT. (INTEGRAL PROTEIN ISOLATION??) THE EXAMPLE IS CYTOCHROME C. IT IS ONE OF THE PROTEINS THAT IS INVOLVED IN OXIDATION- REDUCTIONS IN THE OX-PHOS PATHWAY. IT IS ATTACHED TO THE CRISTAE ON THE INTERMEMBRANE SIDE OF MITO- CHONDRIA.
HOW ARE LIPIDS & PROTEINS “ASSOCIATED” IN MEMBRANES? IT WAS KNOWN EARLY ON THAT EXTRINSIC MEMBRANE PROTEINS COULD BE EASILY REMOVED FROM MEMBRANES BY EXPOSING THE MEMBRANE TO SALT SOLUTIONS THAT WERE STRONGER THAN THE IONIC (SALT) SURFACE FORCES ON THE INNER AND OUTER FACES OF THE LIPIDS THEMSELVES. THESE “FORCES” ONLY SERVE THE ROLE OF KEEPING THE PROTEINS IN PLACE IN A SOMEWHAT LOOSE MANNER. THE EXTRINSIC PROTEINS ATTACH TO THE MEMBRANES BY: IONIC INTERACTIONS WITH THE LIPIDS OR EVEN PARTS OF INTEGRAL MEMBRANE PROTEINS THAT PROJECT OUT OF THE MEMBRANE. EVEN SHORT LOOPS OF HYDROPHOBIC AMINO ACIDS THAT STICK INTO THE LIPID ARE FOUND, BUT THESE ARE NOT SO PERMANENT THAT THEY CAN’T BE REMOVED BY “SALTING OUT” THE OTHER PARTS OF THE PROTEIN.
THE ATTACHMENTS OF INTEGRAL MEMBRANE PROTEINS WITHIN THE LIPID PORTION OF THE MEMBRANE ARE MORE ROBUST (STRONGER) THAN PERIPHERAL PROTEINS AND ARE OFTEN ASSOCIATED WITH SOME MEMBRANE FUNCTION . IT HAS ALREADY BEEN SHOWN THAT a-HELICES AND b-STRUCTURES (i. e., BARRELS AND STRANDS) OF INTEGRAL MEMBRANE PROTEINS ARE USED JUST TO HAVE A PROTEIN CROSS (PENETRATE) THE LIPID MEMBRANE. IN ADDITION: ANCHORING (= POSITIONING) OF INTEGRAL PROTEINS TO MEMBRANE LIPIDS MAY ALSO REQUIRE OR SUBSTITUTE SEPARATE KINDS OF BONDS FOR PROTEIN FUNCTION TO OCCUR – AMIDE-LINKED MYRISTOYL ANCHORS; THIOESTER-LINKED FATTY ACYL ANCHORS; THIOETHER-LINKED PRENYL ANCHORS; AND GLYCOSYL PHOSPHATIDYLINOSITOL ANCHORS. MYRISTOYL = PROTEIN LINKED TO MYRISTIC ACID (C14 FATTY ACID) FATTY ACYL = PROTEIN LINKED TO A FATTY ACID (NON-SPECIFIC) PRENYL = PROTEIN LINKED TO A PRENYL GROUP: C-C=C-C-(CH3)2
HERE ARE TWO EXAMPLES: THE PROTEIN ON THE LEFT IS LINKED TO MYRISTIC ACID THROUGH AN AMIDE GROUP. THE MYRISTIC ACID, IN TURN IS PART OF THE LIPID BI-LAYER. THE PROTEIN ON THE RIGHT IS LINKED TO PALMITIC ACID THROUGH A THIOESTER. THE PALMITIC ACID IS PART OF THE LIPID BI-LAYER. HOWEVER, IN ADDITION, THE PROTEIN TRAVERSES THE LIPID BILAYER WITH SEVERAL a-HELICES. PALMITIC ACID MYRISTIC ACID PROTEIN PROTEIN
A REAL EXAMPLE OF AN INTRINSIC PROTEIN WHOSE POLYPEPTIDE CHAINS DO NOT ENTER THE BILIPID LAYER IS THE “REGGIE” PROTEIN. REGGIE-1 IS HELD TO ITS MEMBRANE BY MYRISTOL- ATION AND PALMITOLATION NEAR THE N-TERMINAL END OF THE PROTEIN. THESE DOUBLE ANCHORS DIRECT THE REMAINDER OF THE MOLECULE TO CARRY OUT ITS FUNCTIONS ON THE INSIDE OF THE CELL. REGGIE-1 HAS BEEN ASSOCIATED WITH THE REFORMATION OF THE CYTOSKELATIN, CELL ADHESION, ENDOCYTOSIS AND OTHER FUNCTIONS. OUTSIDE OF CELL
THE CONCEPT OF A MEMBRANE “RAFT” IT IS CURRENTLY HYPOTHESIZED (=THOUGHT) THAT THERE ARE REGIONS ON A MEMBRANE BILIPID IN WHICH LATERAL DIFFUSION (=MOVEMENT) IS HINDERED. THESE REGIONS ARE DESCRIBED AS BEING RICH IN CHOLESTEROL AND GLYCOSPHINGOLIPIDS (HAVING ACYL GROUPS THAT ARE LONG CHAINED AND SATURATED). SUCH AREAS ARE ALSO KNOWN AS MICRODOMAINS OR MEMBRANE RAFTS. THEY ARE CONSIDERED TO BE SMALL, MOBILE AND WITH SHORT ½ LIVES. ONLY INDIRECT EVIDENCE HAS POINTED TO THEIR EXISTENCE. THE ADVANTAGE OF A RAFT IS TO FACILITATE THE FUNCTIONS OF SIGNALING (=RECEPTOR) PROTEINS. GPI = GLYCOSYL PHOSPHO- INOSITOL ANCHOR. CAVEOLIN = A PROTEIN INVOLVED IN SIGNALING & ENDOCYTOSIS. NOTE: ORANGE GLYCO- SPHINGOLIPIDS AND YELLOW CHOLESTEROL.
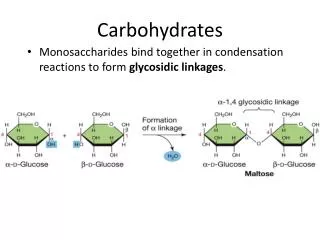
CARBOHYDRATES IN MEMBRANES CARBOHYDRATES ARE, OF COURSE, NOT COMPATIBLE WITH THE APOLAR NATURE OF A MEMBRANE. THEY ARE ASSOCIATED WITH MEMBRANE LIPIDS UNDER THE GROUP KNOWN AS SPHINGOLIPIDS (= GLYCOLIPIDS) AND THEY ARE ALSO COVALENTLY BOUND TO MANY MEMBRANE PROTEINS (= GLYCOPROTEINS). THEY ARE POSITIONED ON THE OUTER FACE OF A MEMBRANE (OFTEN ON THE OUTSIDE). THEY MAY FORM A LOOSE ASSOCIATIONS WITH OTHER CARBOHYDRATES KNOWN COLLECTIVELY AS THE GLYCOCALIX (LITERALLY SUGAR COAT) OF A CELL. MANY CARBOHYDRATES ON MEMBRANES SERVE THE ROLE OF IMMUNE IDENTIFIERS (“FRIEND OR FOE?”) FOR THE ORGANISM. OTHERS MAKE UP COMPLEX SYSTEMS THAT COAT THE MEMBRANES OF MICROOGANISMS AND SERVE AS PROTECTIVE DEVICES FROM ATTACK OR ACT EVEN AS OSMOTIC STABILIZERS FOR THE ORGANISMS’ MEMBRANE. SYNDECAN (RIGHT) ACTS AS A MOLECULAR GLUE FOR OTHER MOLECULES IN THE EXTRA- CELLULAR MATRIX.
A FEW OTHER FACTS ABOUT CARBOHYDRATES • THAT BELONG TO MEMBRANES: • The glycocalyx may be involved in a process called • “lymphocyte homing” that helps guide white blood cells • to a cell requiring immunological intervention – that is • they serve as markers. • -Many of the oligosaccharides that make up a cell surface • glycocalyx end in sialic acid which has a negative charge. • This charge repels substances from approaching a cell • surface.
PUTTING IT ALL TOGETHER MEMBRANES ARE COMPOSED OF LIPIDS, PROTEINS AND CARBOHYDRATES FORMED TO MAXIMIZE THE MEMBRANE’S FUNCTION Remember that carbohydrates only are associated with membranes by binding to certain lipids and proteins. They do not enter the bilipid layer.
WHAT IS ESSENTIAL TO UNDERSTAND: • The nature of a bilipid layer in a cell membrane and how phospholipids • form the double layer. • How does one distinguish between a micelle, a bilipid layer and a liposome? • How do sphingolipids and cholesterol contribute to the characteristics of a • membrane? • What is important about fatty acid chain length and degree of unsaturation • in membrane fluidity? • What is meant by “transition” in a membrane? • How are membranes asymetric and what does that impose on a membrane? • What was the contribution of Davson & Danielli to the understanding of • membrane structure? • How does one distinguish between an intrinsic (integral) and extrinsic • (peripheral) membrane protein? Give an example. • How MIGHT a hydropathy plot identify an intrinsic protein? • How do proteins and lipids associate (bind) together in a membrane? • How do intrinsic proteins bind to a lipid bilayer if they do not penetrate the • the membrane with alpha helices and what are membrane rafts? • 12. How do carbohydrates associate with membrane bilayers?