Download
1 / 1
10 likes | 88 Vues
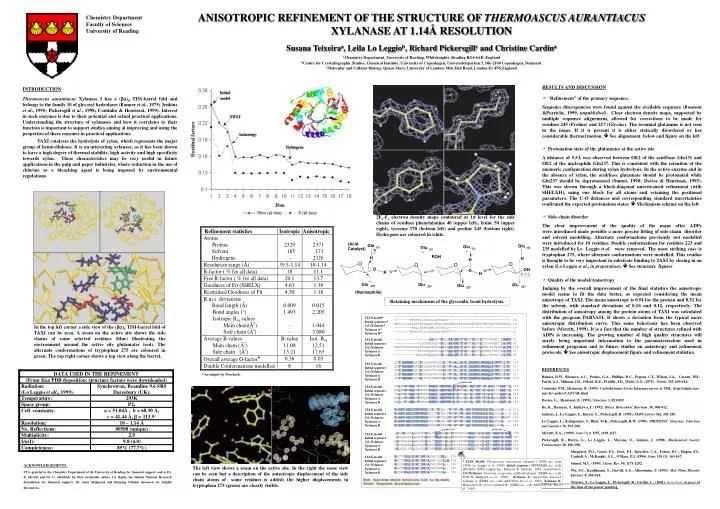
2F o -F c electron density maps contoured at 1 s level for the side chains of residues phenylalanine 40 (upper left), lysine 50 (upper right), tyrosine 170 (bottom left) and proline 245 (bottom right). Hydrogens are coloured in white. 5.5Å.

E N D
2Fo-Fc electron density maps contoured at 1s level for the side chains of residues phenylalanine 40 (upper left), lysine 50 (upper right), tyrosine 170 (bottom left) and proline 245 (bottom right). Hydrogens are coloured in white. 5.5Å 1TAX ------------------------------------------------------------ Q9UQZ4 --------MVRPTILLTSLLLAPFAAAS-------------------------------- 20 FOFCH MHTLSVLLALAPVSALAQAPIWGQCGGNGWTGATTCASGLKCEKINDWYYQCVPGSGGSE 60 AKXYNA ---------MVQIKAAALAMLFASHVLS-------------------------------- 19 MGXYN33A --------MKASSVLLGLAPLAALAAPTP------------------------------- 21 1TAX ------------------ASAQSVDQLIKARGKVYFGVATDQNRLTTG-KNAAIIQANFG 41 Q9UQZ4 ------------PILEERQAAQSVDQLIKARGKVYFGVATDQNRLTTG-KNAAIIQADFG 67 FOFCH PQPSSTQGGGTPQPTGGNSGGTGLDAKFKAKGKQYFGTEIDHYHLNNN-PLINIVKAQFG 119 AKXYNA ------------EPIEPRQASVSIDSKFKAHGKKYLGNIGDQYTLTKNSKTPAVIKADFG 67 MGXYN33A -----------EAELSARQAQQSIDALMKAKGKLYFGTATDQGLLNTG-KNSAIIKADFG 69 1TAX QVTPENSMKWDATEPSQGNFNFAGADYLVNWAQQNGKLIRGHTLVWHSQLPSWVTSITDK 101 Q9UQZ4 QVTPENSMKWDATEPSQGNFNFAGADYLVNWAQQNGKLIRGHTLVWHSQLPSWVSSITDK 127 FOFCH QVTCENSMKWDAIEPSRNSFTFSNADKVVDFATQNGKLIRGHTLLWHSQLPQWVQNINDR 179 AKXYNA ALTPENSMKWDATEPSRGQFSFSGSDYLVNFAQSNNKLIRGHTLVWHSQLPSWVQAITDK 127 MGXYN33A QVTPENSMKCQSLENTRGQYNWAPADALVNFAVSNNKSIRGHTLIWHSQLPGWVNNINDR 129 1TAX NTLTNVMKNHITTLMTRYKGKIRAWDVVN-EAFNQNGSLRSTVFLNVIGEDYIPIAFQTA 160 Q9UQZ4 NTLTNVMKNHITTLMTRYKGKIRAWDVVN-EAFNEDGSLRQTVFLNVIGEDYIPIAFQTA 186 FOFCH STLTAVIENHVKTMVTRYKGKILQWDVVNNEIFAEDGNLRDSVFSRVLGEDFVGIAFRAA 239 AKXYNA NTLIEVMKNHITTVMQHYKGKIYAWDVVN-EIFNEDGSLRDSVFYKVIGDDYVRIAFETA 186 MGXYN33A NQLTTVIQNHVATVMGRWKGKIRAWDVVN-EIFNEDGTMRQSVFSRVLGEDFVRIAFEAA 188 1TAX RAADPNAKLYINDYNLDSATYPKTQA-IVNRVKQWRAAGVPIDGIGSQTHLSAGQG---A 216 Q9UQZ4 RAADPNAKLYINDYNLDSASYPKTQA-IVNRVKQWRAAGVPIDGIGSQTHLSAGQG---A 242 FOFCH RAADPAAKLYINDYNLDKSDYAKVTRGMVAHVNKWIAAGIPIDGIGSQGHLAAPSGWNPA 299 AKXYNA RAADPNAKLYINDYNLDSASYPKLAG-MVSHVKKWIEAGIPIDGIGSQTHLSAGGG---A 242 MGXYN33A RKADPNAKLYINDYNLDRPNAGKLTKGMVGHVKKWVGAGVPIDGIGRQGHLQSGQG---- 244 1TAX -GVLNALPLLASAGTPEVAITELDVAGASPTDYVNVVNACLNVSSCVGITVWGVADPDSW 275 Q9UQZ4 -SVLQALPLLASAGTPEVAITELDVAGASSTDYVNVVNACLNVQSCVGITVWGVADPDSW 301 FOFCH SGVPAALRALAASDAKEIAITELDIAGASANDYLTVMNACLAVPKCVGITVWGVSDKDSW 359 AKXYNA -GISGALNALAGAGTKEIAVTELDIAGASSTDYVEVVEACLDQPKCIGITVWGVADPDSW 301 MGXYN33A NGLGQGIKGLGDSGVKEVGGNELDIQGNNGNEFGGGNKACLPVPACVGIPAWGVRDNDSW 304 1TAX RASTTPLLFDGNFNPKPAYNAIVQNLQ- 302 Q9UQZ4 RASTTPLLFDGNFNPKPAYNAIVQDLQQ 329 FOFCH RPGDNPLLYDSNYQPKAAFNALANAL-- 385 AKXYNA RSSSTPLLFDSNYNPKPAYTAIANAL-- 327 MGXYN33A RPQGNPLLFDSNYNPKPAYNSVVQALK- 331 Red: Residues where mutations had to be made Green: Sequence discrepancies 1TAX model Initial sequence Cel./Xylanase Xylanase A Xylanase B 1TAX model Initial sequence Cel./Xylanase Xylanase A Xylanase B In the top left corner a side view of the (ba)8 TIM-barrel fold of TAXI can be seen. A zoom on the active site shows the side chains of some selected residues (blue) illustrating the environment around the active site glutamates (red). The alternate conformations of tryptophan 275 are coloured in green. The top right corner shows a top view along the barrel. 1TAX model Initial sequence Cel./Xylanase Xylanase A Xylanase B Active site Active site 1TAX model Initial sequence Cel./Xylanase Xylanase A Xylanase B 1TAX model Initial sequence Cel./Xylanase Xylanase A Xylanase B Active site 1TAX model Initial sequence Cel./Xylanase Xylanase A Xylanase B The left view shows a zoom on the active site. In the right the same view can be seen but a description of the anisotropic displacement of the side chain atoms of some residues is added: the higher displacements in tryptophan 275 (green) are clearly visible. 1TAX model Initial sequence Cel./Xylanase Xylanase A Xylanase B 1TAX Model: Thermoascus aurantiacus xylanase I (PDB acc. code 1TAX; Lo Leggio et al., 1999). Initial sequence: SPTREMBL acc. code Q9UQZ4 (DNA sequencing; Bousson & Parriche, 1999, unpublished). Cel./Xylanase: Fusarium oxysporum cellulase/xylanase (EMBL acc. code FOFCH; Sheppard et al., 1994). Xylanase A: Aspergillus kawachii xylanase A (EMBL acc. code AKXYNA; Ito et al., 1992). Xylanase B: Magnatoporthe grisea xylanase B ( EMBL acc. code MGXYN33A; Wu et al., 1995). As output by Procheck. ANISOTROPIC REFINEMENT OF THE STRUCTURE OF THERMOASCUS AURANTIACUS XYLANASE AT 1.14Å RESOLUTION Susana Teixeiraa, Leila Lo Leggiob, Richard Pickersgillc and Christine Cardina aChemistry Department, University of Reading, Whiteknights, Reading RG6 6AD, England bCentre for Crystallographic Studies, Chemical Institute, University of Copenhagen, Universitetsparken 5, DK-2100 Copenhagen, Denmark cMolecular and Cellular Biology, Queen Mary, University of London, Mile End Road, London E1 4NS,England Chemistry Department Faculty of Sciences University of Reading • RESULTS AND DISCUSSION • “Refinement” of the primary sequence. Sequence discrepancies were found against the available sequence (Bousson &Parriche, 1999, unpublished). Clear electron density maps, supported by multiple sequence alignement, allowed for corrections to be made for residues 245 (Proline) and 217 (Glycine). The terminal glutamine is not seen in the maps. If it is present it is either statically disordered or has considerable thermal motion. See alignement below and figure on the left • Protonation state of the glutamates at the active site A distance of 5.5Å was observed between OE2 of the acid/base Glu131 and OE2 of the nucleophile Glu237. This is consistent with the retention of the anomeric configuration during xylan hydrolysis. In the active enzyme and in the absence of xylan, the acid/base glutamate should be protonated while Glu237 should be deprotonated (Sinnot, 1990; Davies & Henrissat, 1995). This was shown through a block-diagonal unrestrained refinement (with SHELXH), using one block for all atoms and retaining the positional parameters. The C-O distances and corresponding standard uncertainties confirmed the expected protonation states Mechanism scheme on the left • Side-chain disorder The clear improvement of the quality of the maps after ADPs were introduced made possible a more precise fitting of side-chain disorder and solvent modelling. Alternate conformations previously not modelled were introduced for 10 residues. Double conformations for residues 223 and 235 modelled by Lo Leggio et al. were removed. The most striking case is tryptophan 275, where alternate conformations were modelled. This residue is thought to be very important in substrate binding to TAXI by closing in on xylan (Lo Leggio et al., in preparation). See structure figures • Quality of the model/Anisotropy Judging by the overall improvement of the final statistics the anisotropic model seems to fit the data better, as expected considering the mean anisotropy of TAXI. The mean anisotropy is 0.54 for the protein and 0.52 for the solvent, with standard deviations of 0.16 and 0.12, respectively. The distribution of anisotropy among the protein atoms of TAXI was calculated with the program PARVATI. It shows a deviation from the typical more anisotropic distribution curve. This same behaviour has been observed before (Merritt, 1999). It is a fact that the number of structures refined with ADPs is increasing. The growing number of high quality structures will surely bring important information to the parameterisation used in refinement programs and to future studies on anisotropy and refinement protocols. See anisotropic displacement figure and refinement statistics. INTRODUCTION Thermoascus aurantiacus Xylanase I has a (ba)8 TIM-barrel fold and belongs to the family 10 of glycosyl hydrolases (Banner et al., 1975; Jenkins et al., 1995; Pickersgill et al., 1998; Coutinho & Henrissat, 1999). Interest in such enzymes is due to their potential and actual practical applications. Understanding the structure of xylanases and how it correlates to their function is important to support studies aiming at improving and using the properties of these enzymes in practical applications. TAXI catalyses the hydrolysis of xylan, which represents the major group of hemicelluloses. It is an interesting xylanase, as it has been shown to have a high degree of thermal stability, high activity and high specificity towards xylan. These characteristics may be very useful in future applications in the pulp and paper industries, where reduction in the use of chlorine as a bleaching agent is being imposed by environmental regulations. Retaining mechanism of the glycosidic bond hydrolysis. REFERENCES Banner, D.W., Bloomer, A.C., Petsko, G.A., Phillips, D.C., Pogson, C.I., Wilson, I.A., Corran, P.H., Furth, A.J., Milman, J.D., Offord, R.E., Priddle, J.D., Waley, S.G. (1975). Nature 255, 609-614. Coutinho, P.M., Henrissat, B. (1999). Carbohydrate-Active Enzymes server at URL: http://afmb.cnrs-mrs.fr/~pedro/CAZY/db.html Davies, G., Henrissat, B. (1995). Structure 3, 853-859. Ito, K., Ikemasu, T., Ishikawa, T. (1992). Biosci. Biotechnol. Biochem. 56, 906-912. Jenkins, J., Lo Leggio, L., Harris, G., Pickersgill, R. (1995). FEBS Letters 362, 281-285. Lo Leggio, L., Kalogiannis, S., Bhat, M.K., Pickersgill, R.W. (1999). PROTEINS: Structure, Function and Genetics 36, 295-306. Merritt, E.A., (1999). Acta Cryst. D55, 1109-1117. Pickersgill, R., Harris, G., Lo Leggio, L., Mayans, O., Jenkins, J. (1998). Biochemical Society Transactions 26, 190-198. Sheppard, P.O., Grant, F.J., Oort, P.J., Sprecher, C.A., Foster, D.C., Hagen, F.S., Upshall, A., McKnight, G.L., O'Hara, P.J. (1994). Gene 150 (1), 163-167. Sinnot, M.L. (1990). Chem. Rev. 90, 1171-1202. Wu, S.C., Kauffmann, S., Darvill, A.G., Albersheim, P. (1995). Mol. Plant Microbe Interact. 8, 506-514. Teixeira, S., Lo Leggio, L., Pickersgill, R., Cardin, C., (2001). Acta Cryst, in press at the time of the poster printing. ACKNOWLEDGEMENTS: ST is grateful to the Chemistry Department of the University of Reading for financial support, and to Dr. E. Merritt and Dr. G. Sheldrick for their invaluable advice. LL thanks the Danish National Research Foundation for financial support, Dr. Anne Mølgaard and Henning Osholm Sørensen for helpful discussions.