Download

1 / 52
530 likes | 650 Vues
The Ribosome Flow Model. Michael Margaliot School of Elec. Eng. Tel Aviv University, Israel. Tamir Tuller (Tel Aviv University) Eduardo D. Sontag (Rutgers University). Joint work with: . Overview. Ribosome flow Mathematical models: from TASEP to the Ribosome Flow Model (RFM)

E N D
The Ribosome Flow Model Michael Margaliot School of Elec. Eng. Tel Aviv University, Israel Tamir Tuller (Tel Aviv University) Eduardo D. Sontag (Rutgers University) Joint work with:
Overview Ribosome flow Mathematical models: from TASEP to the Ribosome Flow Model (RFM) Analysis of the RFM+biological implications: • Contraction (after a short time) • Monotone systems • Continued fractions
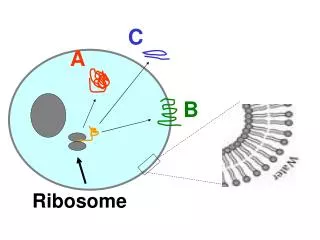
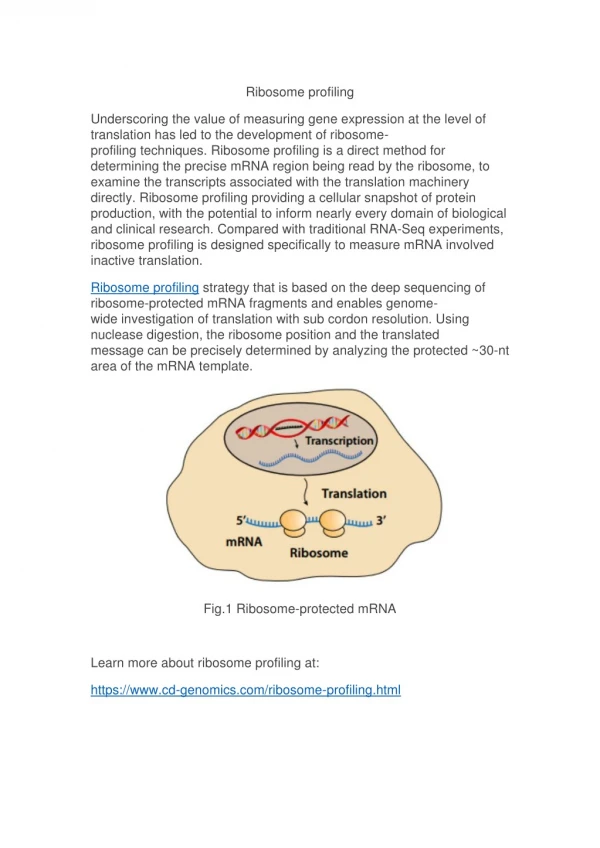
From DNA to Proteins • Transcription: the cell’s machinery • copies the DNA into mRNA • The mRNA travels from the nucleus to • the cytoplasm • Translation: ribosomes “read” the mRNA and produce a corresponding chain of amino-acids
Translation http://www.youtube.com/watch?v=TfYf_rPWUdY http://www.youtube.com/watch?v=TfYf_rPWUdY
Ribosome Flow • During translation several ribosomes • read the same mRNA. Ribosomes • follow each other like cars traveling • along a road. • Mathematical models for ribosome • flow: TASEP* and the RFM. *Zia, Dong, Schmittmann, “Modeling Translation in Protein Synthesis with TASEP: A Tutorial and Recent Developments”, J. Statistical Physics, 2011
Totally Asymmetric Simple Exclusion Process (TASEP) A stochastic model: particles hop along a lattice of consecutive sites Movement is unidirectional (TA) Particles can only hop to emptysites (SE)
Simulating TASEP At each time step, all the particles are scanned and hop with probability , if the consecutive site is empty. This is continued until steady state.
Analysis of TASEP* Mean field approximations Bethe ansatz *Schadschneider, Chowdhury & Nishinari, Stochastic Transport in Complex Systems: From Molecules to Vehicles, 2010. 8
Ribosome Flow Model* A deterministic model for ribosome flow. mRNA is coarse-grained into consecutive sites. Ribosomes reach site 1 with rate , but can only bind if the site is empty. *Reuveni, Meilijson, Kupiec, Ruppin & Tuller, “Genome-scale analysis of translation elongation with a ribosome flow model”, PLoSComput. Biol., 2011
Ribosome Flow Model State-variables: (normalized) number of ribosomes at site i Parameters: >0initiation rate >0 transition rates between consecutive sites
Ribosome Flow Model Just like TASEP, this encapsulates both unidirectional movement and simple exclusion.
Simulation Results e All trajectories emanating from remain in , and converge to a unique equilibrium point e.
Analysis of the RFM Uses tools from: • Contraction theory • Monotone systems theory • Analytic theory of continued fractions
Contraction Theory* The system: • is contracting on a convex set K, with • contraction rate c>0, if • for all • *Lohmiller & Slotine, “On Contraction Analysis • for Nonlinear Systems”, Automatica, 1988.
Contraction Theory Trajectories contract to each other at an exponential rate. x(t,0,a) a x(t,0,b) b
Implications of Contraction 1. Trajectories converge to a unique equilibrium point; 2. The system entrains to periodic excitations.
Contraction and Entrainment* Definition is T-periodic if Theorem The contracting and T-periodic • system admits a unique • periodic solution of period T, and • *Russo, di Bernardo, Sontag, “Global Entrainment of Transcriptional Systems to Periodic Inputs”, PLoSComput. Biol., 2010.
How to Prove Contraction? The Jacobian of is the nxn matrix
How to Prove Contraction? The infinitesimal distance between trajectories evolves according to This suggests that in order to prove contraction we need to (uniformly) bound J(x).
How to Prove Contraction? Let be a vector norm. The induced matrix norm is: The induced matrix measure is:
How to Prove Contraction? Intuition on the matrix measure: Consider Then to 1st order in so
Proving Contraction Theorem Consider the system for all then the If • system is contracting on K with contraction • rate c. Comment 1: all this works for Comment 2: is Hurwitz.
Application to the RFM For n=3, and for the matrix measure induced by the L1 vector norm: for all The RFM is on the “verge of contraction.”
RFM is not Contracting on C For n=3: • so for is singular and thus not Hurwitz.
Contraction After a Short Transient (CAST)* Definition is a CAST if • there exists such that -> Contraction after an arbitrarily small transient in time and amplitude. • *M., Sontag & Tuller, “Entrainment to Periodic Initiation and Transition Rates in the Ribosome Flow Model”, submitted, 2013.
Motivation for Contraction after a Short Transient (CAST) Contraction is used to prove asymptotic properties (convergence to equilibrium point; entrainment to a periodic excitation).
Application to the RFM Theorem The RFM is CAST on . Corollary 1 All trajectories converge to a unique equilibrium point e.* Biological interpretation: the parameters determine a unique steady-state of ribosome distributions and synthesis rate; not affected by perturbations. • *M.& Tuller, “Stability Analysis of the Ribosome Flow Model”, IEEE TCBB, 2012.
Application to the RFM Theorem The RFM is CAST on C. Corollary 2 Trajectories entrain to periodic initiation and/or transition rates (with a common period T).* Biological interpretation: ribosome distributions and synthesis rate converge to a periodic pattern, with period T. • *M., Sontag & Tuller, “Entrainment to Periodic Initiation and Transition Rates in the Ribosome Flow Model”, submitted, 2013.
Entrainment in the RFM Here n=3,
Analysis of the RFM Uses tools from: • Contraction theory • Monotone systems theory • Analytic theory of continued fractions
Monotone Dynamical Systems* Define a (partial) ordering between vectors in Rn by: Definitionis called monotone if i.e., the dynamics preserves the partial ordering. *Smith, Monotone Dynamical Systems: An Introduction to the Theory of Competitive and Cooperative Systems, AMS, 1995
Monotone Dynamical Systems in the Life Sciences Used for modeling a variety of biochemical networks:* • behavior is ordered and robust with respect to parameter values • large systems may be modeled as interconnections of monotone subsystems. *Sontag, “Monotone and Near-Monotone Biochemical Networks”, Systems & Synthetic Biology, 2007
When is a System Monotone? Theorem (Kamke Condition.) Suppose that f satisfies: then is monotone. Intuition: assume monotonicityislost, then
Verifying the Kamke Condition Theorem cooperativityKamke condition ( system is monotone) • Definition is calledcooperative • if • This means that increasing increases
Application to the RFM • Proposition The RFM is monotone on C. Every off-diagonal entry is non- negative on C. Thus, the RFM is a cooperative system. • Proof:
RFM is Cooperative • Intuition if x2 increases then increase. A “traffic jam” in a site induces “traffic jams” in the neighboring sites. and
RFM is Monotone • Biological implication: a larger initial • distribution of ribosomes induces a • larger distribution of ribosomes for all • time.
Analysis of the RFM Uses tools from: • Contraction theory • Monotone systems theory • Analytic theory of continued fractions
Continued Fractions Suppose (for simplicity) that n =3. Then Let denote the unique equilibrium point in C. Then
Continued Fractions . . This yields: Every eican be expressed as a continued fraction of e3 .
Continued Fractions . Furthermore, e3 satisfies: . . . This is a second-order polynomial equation in e3. In general, this is a th–order polynomial equation in en.
Homogeneous RFM In certain cases, all the transition rates are approximately equal.* In the RFM this can be modeled by assuming that This yields the Homogeneous Ribosome Flow Model (HRFM). Analysis is simplified because there are only two parameters. *Ingolia, Lareau & Weissman, “Ribosome Profiling of Mouse Embryonic Stem Cells Reveals the Complexity and Dynamics of Mammalian Proteomes”, Cell, 2011
HRFM and Periodic Continued Fractions In the HRFM, This is a periodic continued fraction, and we can say a lot more about e.
Equilibrium Point in the HRFM* Theorem In the HRFM, Biological interpretation: This provides an explicit expression for the capacity of a gene. *M. & Tuller, “On the Steady-State Distribution in the Homogeneous Ribosome Flow Model”, IEEE TCBB, 2012
mRNA Circularization* *Craig, Haghighat, Yu & Sonenberg, ”Interaction of Polyadenylate-Binding Protein with the eIF4G homologue PAIP enhances translation”, Nature, 1998
RFM as a Control System This can be modeled by the RFM with Input and Output (RFMIO): and then closing the loop via Remark: The RFMIO is a monotone control system.* *Angeli & Sontag, “Monotone Control Systems”, IEEE TAC, 2003
RFM with Feedback* • Theorem The closed-loop system admits • an equilibrium point e that is globally • attracting in C. • Biological implication: as before, but this • is probably a better model for translation • in eukaryotes. *M. & Tuller, “Ribosome Flow Model with Feedback”, J. Royal Society -Interface, to appear
RFM with Feedback* • Theorem In the homogeneous case, • where • Biological implication: may be useful, • perhaps, for re-engineering gene translation.