Download
1 / 1
10 likes | 123 Vues
Glia in Cell Culture Insert Support Neurons in vitro.

E N D
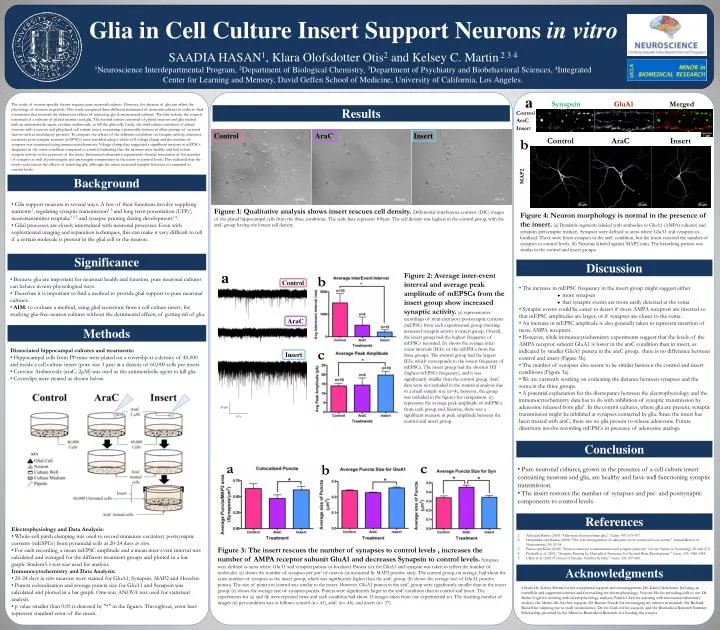
Glia in Cell Culture Insert Support Neurons in vitro SAADIA HASAN1, Klara Olofsdotter Otis2 and Kelsey C. Martin 2 3 41Neuroscience Interdepartmental Program, 2Department of Biological Chemistry, 3Department of Psychiatry and Biobehavioral Sciences, 4Integrated Center for Learning and Memory, David Geffen School of Medicine, University of California, Los Angeles. a Synapsin GluA1 Merged The study of neuron-specific factors requires pure neuronal cultures. However, the absence of glia can affect the physiology of neurons negatively. This study compared three different treatments of neuronal cultures in order to find a treatment that reversed the deleterious effects of removing glia from neuronal cultures. The first culture, the control, consisted of a mixture of plated neurons and glia. The second culture consisted of plated neurons and glia treated with an antimetabolic agent, cytosine arabinoside, to kill the glial cells. Lastly, the third culture consisted of plated neurons with a neuron and glia plated cell culture insert, containing a permeable bottom to allow passage of secreted factors such as modulatory proteins. To compare the effects of the different conditions on synaptic activity, miniature excitatory post-synaptic currents (mEPSCs) were recorded using a whole-cell voltage clamp and the number of synapses was monitored using immunocytochemistry. Voltage-clamp data suggested a significant increase in mEPSCs frequency in the insert condition compared to control, indicating that the neurons were healthy and had robust synaptic activity in the presence of the insert. Immunocytochemistry experiments showed restoration of the number of synapses as well as postsynaptic and presynaptic components in the insert to control levels. This indicated that the insert could rescue the effects of removing glia, although the insert increased synaptic function as compared to control levels. Results Control AraC Insert Control AraC Insert b Control AraC Insert MAP2 Background • Glia support neurons in several ways. A few of their functions involve supplying nutrients1, regulating synaptic transmission1 3 and long term potentiation (LTP)3, neurotransmitter reuptake1 3 5 and synapse pruning during development1 4. • Glial processes are closely intertwined with neuronal processes. Even with sophisticated imaging and separation techniques, this can make it very difficult to tell if a certain molecule is present in the glial cell or the neuron. Figure 1: Qualitative analysis shows insert rescues cell density. Differential interference contrast (DIC) images of the plated hippocampal cells from the three conditions. The scale bars represent 100μm. The cell density was highest in the control group, with the araC group having the lowest cell density. Figure 4: Neuron morphology is normal in the presence of the insert. (a) Dendritic segments labeled with antibodies to GluA1 (AMPAr subunit) and synapsin (presynaptic marker). Synapses were defined as areas where GluA1 and synapsin co-localized. There were fewer synapses in the araC condition, but the insert restored the number of synapses to control levels. (b) Neurons labeled against MAP2 stain. The branching pattern was similar in the control and insert groups. Significance Discussion a Figure 2: Average inter-event interval and average peak amplitude of mEPSCs from the insert group show increased synaptic activity. (a) representative recordings of mini-excitatory postsynaptic currents (mEPSC) from each experimental group showing increased synaptic activity in insert group. Overall, the insert group had the highest frequency of mEPSCs recorded. (b) shows the average inter-event intervals (IEIs) of the mEPSCs from the three groups. The control group had the largest IEIs, which corresponds to the lowest frequency of mEPSCs. The insert group had the shortest IEI (highest mEPSCs frequency), and it was significantly smaller than the control group. AraC data were not included in the statistical analysis due to a small sample size (n=4), however, the group was included in the figures for comparison. (c) represents the average peak amplitude of mEPSCs from each group and, likewise, there was a significant increase in peak amplitude between the control and insert group. b • • Because glia are important for neuronal health and function, pure neuronal cultures can behave in non-physiological ways. • Therefore it is important to find a method to provide glial support to pure neuronal cultures. • AIM: to evaluate a method, using glial secretions from a cell culture insert, for studying glia-free neuron cultures without the detrimental effects of getting rid of glia. Control • The increase in mEPSC frequency in the insert group might suggest either • more synapses • that synaptic events are more easily detected at the soma • Synaptic events could be easier to detect if more AMPA receptors are inserted so that mEPSC amplitudes are larger, or if synapses are closer to the soma. • An increase in mEPSC amplitude is also generally taken to represent insertion of more AMPA receptors. • However, while immunocytochemistry experiments suggest that the levels of the AMPA receptor subunit GluA1 is lower in the araC condition than in insert, as indicated by smaller GluA1 puncta in the araC group, there is no difference between control and insert (Figure 3b). • The number of synapses also seems to be similar between the control and insert conditions (Figure 3a). • We are currently working on evaluating the distance between synapses and the soma in the three groups. • A potential explanation for the discrepancy between the electrophysiology and the immunocytochemistry data has to do with inhibition of synaptic transmission by adenosine released from glia2. In the control cultures, where glia are present, synaptic transmission might be inhibited at synapses contacted by glia. Since the insert has been treated with araC, there are no glia present to release adenosine. Future directions involve recording mEPSCs in presence of adenosine analogs. AraC Methods c • Dissociated hippocampal cultures and treatments: • Hippocampal cells from P0 mice were plated on a coverslip at a density of 40,000 and inside a cell culture insert (pore size 3 μm) at a density of 60,000 cells per insert. • Cytosine Arabinoside (araC; 2μM) was used as the antimetabolic agent to kill glia. • Coverslips were treated as shown below. Insert Conclusion a c b • Pure neuronal cultures, grown in the presence of a cell culture insert containing neurons and glia, are healthy and have well functioning synaptic transmission. • The insert restores the number of synapses and pre- and postsynaptic components to control levels. References • Electrophysiology and Data Analysis: • Whole-cell patch clamping was used to record miniature excitatory postsynaptic currents (mESPCs) from pyramidal cells at 20-24 days in vitro. • For each recording, a mean mEPSC amplitude and a mean inter-event interval was calculated and averaged for the different treatment groups and plotted in a bar graph. Student’s t-test was used for analysis. • Immunocytochemistry and Data Analysis: • 20-24 days in vitro neurons were stained for GluA1, Synapsin, MAP2 and Hoechst. • Puncta colocalization and average puncta size for GluA1 and Synapsin was calculated and plotted in a bar graph. One-way ANOVA was used for statistical analysis. • p value smaller than 0.05 is denoted by "*" in the figures. Throughout, error bars represent standard error of the mean. Allen and Barres (2009) “Glia-more than just brain glue.” Nature, 457: 675-677. Dunwiddie and Masino (2001) "The role and regulation of adenosine in the central nervous system." Annual Review of Neuroscience, 24: 31-55. Paixao and Klein (2010) “Neuron-astrocyte communication and synaptic plasticity.” Current Opinion in Neurobiology, 20: 466-473. Paolicelli et al. (2011) “Synaptic Pruning by Microglia is Necessary for Normal Brain Development.” Science, 333: 1456-1458. Ullian et al. (2001)“Control of Synapse Number by Glia.” Science, 291: 657-660. Figure 3: The insert rescues the number of synapses to control levels , increases the number of AMPA receptor subunit GluA1 and decreases Synapsin to control levels. Synapses were defined as areas where GluA1 and synapsin puncta co-localized. Puncta size for GluA1 and synapsin was taken to reflect the number of molecules. (a) shows the number of synapses per μm2 of neuron (as measured by MAP2 positive area). The control group, on average, had about the same number of synapses as the insert group, which was significantly higher than the araC group. (b) shows the average size of GluA1 positive puncta. The size of puncta in control was similar to the insert. However, GluA1 puncta in the araC group were significantly smaller than in the insert group. (c) shows the average size of synapsin puncta. Puncta were significantly larger in the araC condition than in control and insert. The experiments for (a) and (b) were repeated twice and each condition had about 15 images taken from one experimental set. The resulting number of images (n) per condition was as follows: control (n= 41), araC (n= 46), and insert (n= 37). Acknowledgments I thank Dr. Kelsey Martin for her exceptional support and encouragement, Dr. Klara Olofsdotter for being an incredible and supportive mentor and for teaching me electrophysiology, Victoria Ho for providing cells to use, Dr. Besim Uzgil for assisting with electrophysiology analysis, Patrick Chen for assisting with immunocytochemistry analysis, the Martin lab for their support, Dr. Roman Ferede for encouraging my interest in research, Dr. Richard Russell for inspiring me to study neuroscience, Dr. Ira Clark for his support, and the Biomedical Research Summer Scholarship, provided by the Minor in Biomedical Research, for funding this project.