Download
1 / 1
30 likes | 131 Vues
N15. C. N. Intracellular Notch. Ligand Binding Domain. B. C. A. C4. C9. C22. Ca 2+. C27. D33. D30. N-term. C34. C18. C-term.

E N D
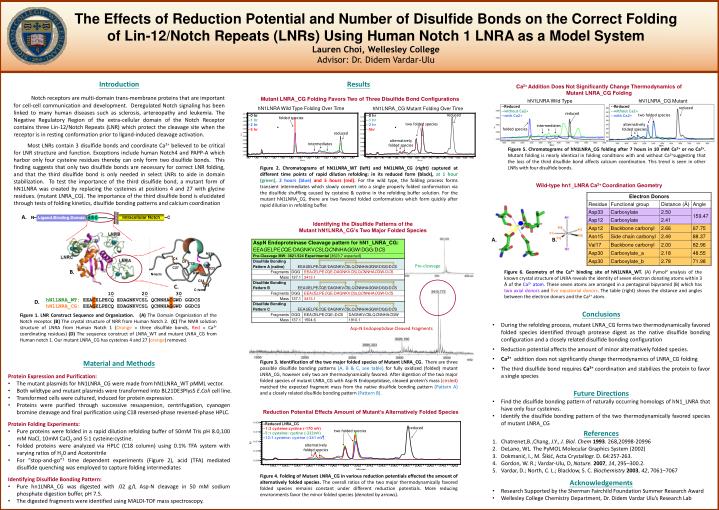
N15 C N Intracellular Notch Ligand Binding Domain B C A C4 C9 C22 Ca2+ C27 D33 D30 N-term C34 C18 C-term The Effects of Reduction Potential and Number of Disulfide Bonds on the Correct Folding of Lin-12/Notch Repeats (LNRs) Using Human Notch 1 LNRA as a Model System Lauren Choi, Wellesley College Advisor: Dr. Didem Vardar-Ulu Introduction Notch receptors are multi-domain trans-membrane proteins that are important for cell-cell communication and development. Deregulated Notch signaling has been linked to many human diseases such as sclerosis, artereopathy and leukemia. The Negative Regulatory Region of the extra-cellular domain of the Notch Receptor contains three Lin-12/Notch Repeats (LNR) which protect the cleavage site when the receptor is in resting conformation prior to ligand-induced cleavage activation. Most LNRs contain 3 disulfide bonds and coordinate Ca2+ believed to be critical for LNR structure and function. Exceptions include human Notch4 and PAPP-A which harbor only four cysteine residues thereby can only form two disulfide bonds. This finding suggests that only two disulfide bonds are necessary for correct LNR folding, and that the third disulfide bond is only needed in select LNRs to aide in domain stabilization. To test the importance of the third disulfide bond, a mutant form of hN1LNRA was created by replacing the cysteines at positions 4 and 27 with glycine residues. (mutant LNRA_CG). The importance of the third disulfide bond is elucidated through tests of folding kinetics, disulfide bonding patterns and calcium coordination Results Ca2+ Addition Does Not Significantly Change Thermodynamics of Mutant LNRA_CG Folding Mutant LNRA_CG Folding Favors Two of Three Disulfide Bond Configurations hN1LNRA Wild Type hN1LNRA_CG Mutant --Reduced --without Ca2+ --with Ca2+ --Reduced --without Ca2+ --with Ca2+ hN1LNRA Wild Type Folding Over Time hN1LNRA_CG Mutant Folding Over Time reduced reduced reduced two folded species --0 hr --1 hr --2 hr --5 hr --0 hr --1 hr --2 hr --5hr folded species two folded species alternatively folded species intermediates folded species reduced alternatively folded species intermediates Figure 5. Chromatograms of hN1LNRA_CG folding after 7 hours in 10 mM Ca2+ or no Ca2+. Mutant folding is nearly identical in folding conditions with and without Ca2+suggesting that the loss of the third disulfide bond affects calcium coordination. This trend is seen in other LNRs with four disulfide bonds. Figure 2. Chromatograms of hN1LNRA_WT (left) and hN1LNRA_CG (right) captured at different time points of rapid dilution refolding: in its reduced form (black), at 1 hour (green),2 hours (blue) and 5 hours (red). For the wild type, the folding process forms transient intermediates which slowly convert into a single properly folded conformation via the disulfide shuffling caused by cysteine & cystine in the refolding buffer solution. For the mutant hN1LNRA_CG, there are two favored folded conformations which form quickly after rapid dilution in refolding buffer. Wild-type hn1_LNRA Ca2+ Coordination Geometry A. Identifying the Disulfide Patterns of the Mutant hN1LNRA_CG’s Two Major Folded Species A. B. Pre-cleavage B. Figure 6. Geometry of the Ca2+ binding site of hN1LNRA_WT. (A) Pymol2 analysis of the known crystal structure of LNRA reveals the identity of seven electron donating atoms within 3 Ă of the Ca2+ atom. These seven atoms are arranged in a pentagonal bipyramid (B) which has two axial donors and five equatorial donors. The table (right) shows the distance and angles between the electron donors and the Ca2+ atom. C. 10 20 30hN1LNRA_WT: EEACELPECQ EDAGNKVCSL QCNNHACGWD GGDCS hN1LNRA_CG: EEAGELPECQ EDAGNKVCSL QCNNHAGGWD GGDCS D. • Conclusions • During the refolding process, mutant LNRA_CG forms two thermodynamically favored folded species identified through protease digest as the native disulfide bonding configuration and a closely related disulfide bonding configuration • Reduction potential affects the amount of minor alternatively folded species. • Ca2+addition does not significantly change thermodynamics of LNRA_CG folding • The third disulfide bond requires Ca2+coordination and stabilizes the protein to favor a single species Figure 1. LNR Construct Sequence and Organization.(A) The Domain Organization of the Notch receptor. (B) The crystal structure of NRR from Human Notch 2. (C) The NMR solution structure of LNRA from Human Notch 1 (Orange = three disulfide bonds, Red = Ca2+ coordinating residues) (D) The sequence construct of LNRA_WT and mutant LNRA_CG from Human notch 1. Our mutant LNRA_CG has cysteines 4 and 27 (orange) removed. Asp-N Endopeptidase Cleaved Fragments • Material and Methods • Protein Expression and Purification: • The mutant plasmids for hN1LNRA_CG were made from hN1LNRA_WT pMML vector. • Both wildtype and mutant plasmids were transformed into BL21DE3PlysS E.Coli cell line. • Transformed cells were cultured, induced for protein expression. • Proteins were purified through successive resuspension, centrifugation, cyanogen bromine cleavage and final purification using C18 reversed-phase reversed-phase HPLC. • Protein Folding Experiments: • Pure proteins were folded in a rapid dilution refolding buffer of 50mM Tris pH 8.0,100 mMNaCl, 10mM CaCl2 and 5:1 cysteine:cystine. • Folded proteins were analyzed via HPLC (C18 column) using 0.1% TFA system with varying ratios of H20 and Acetonitrile • For “stop-and-go”1 time dependent experiments (Figure 2), acid (TFA) mediated disulfide quenching was employed to capture folding intermediates • Identifying Disulfide Bonding Pattern: • Pure hn1LNRA_CG was digested with .02 g/L Asp-N cleavage in 50 mM sodium phosphate digestion buffer, pH 7.5. • The digested fragments were identified using MALDI-TOF mass spectroscopy. Figure 3. Identification of the two major folded species of Mutant LNRA_CG. There are three possible disulfide bonding patterns (A, B & C, see table) for fully oxidized (folded) mutant LNRA_CG, however only two are thermodynamically favored. After digestion of the two major folded species of mutant LNRA_CG with Asp-N Endopeptidase, cleaved protein’s mass (circled) matched the expected fragment mass from the native disulfide bonding pattern (Pattern A) and a closely related disulfide bonding pattern (Pattern B). • Future Directions • Find the disulfide bonding pattern of naturally occurring homologs of hN1_LNRA that have only four cysteines. • Identify the disulfide bonding pattern of the two thermodynamically favored species of mutant LNRA_CG Reduction Potential Effects Amount of Mutant’s Alternatively Folded Species --Reduced LNRA_CG --1:3 cysteine:cystine (-170 mV) --5:1 cysteine: cystine (-233mV) --12:1 cysteine: cystine (-241 mV) reduced References Chatrenet,B.,Chang, J.Y., J. Biol. Chem 1993. 268,20998-20996 DeLano, WL. The PyMOL Molecular Graphics System (2002) Dokmanić, I., M. Šikić, ActaCrystallogr. D. 64:257-263. Gordon, W. R.; Vardar-Ulu, D, Nature.2007, 14, 295–300.2. Vardar, D.; North, C. L.; Blacklow, S. C. Biochemistry2003, 42, 7061–7067 two folded species alternatively folded species Figure 4. Folding of Mutant LNRA_CG in various reduction potentials effected the amount of alternatively folded species. The overall ratios of the two major thermodynamically favored folded species remains constant under different reduction potentials. More reducing environments favor the minor folded species (denoted by arrows). • Acknowledgements • Research Supported by the Sherman Fairchild Foundation Summer Research Award • Wellesley College Chemistry Department, Dr. Didem Vardar Ulu’s Research Lab
