Download

1 / 20
210 likes | 450 Vues
Dr. Kornberg’s key contributions to nucleic acid research: Demonstration of the antiparallel orientation of complementary strands. 10/16/2006 Lecture 9: 1. Geiger Counter.

E N D
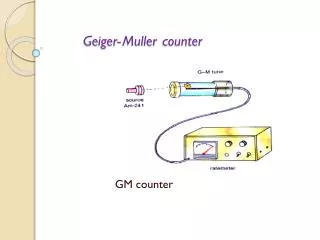
Dr. Kornberg’s key contributions to nucleic acid research: Demonstration of the antiparallel orientation of complementary strands 10/16/2006 Lecture 9: 1 Geiger Counter An ion or electron penetrating the tube (or an electron knocked out of the wall by X-rays or gamma rays) tears electrons off atoms in the gas, and because of the high positive voltage of the central wire, those electrons are then attracted to it. In doing so they gain energy, collide with atoms and release more electrons, until the process snowballs into an "avalanche" which produces an easily detectable pulse of current. With a suitable filling gas, the flow of electricity stops by itself, or else the electrical circuitry can help stop it. The instrument was called a "counter" because every particle passing it produced an identical pulse, allowing particles to be counted (usually electronically) but not telling anything about their identity or energy (except that they must have sufficient energy to penetrate the walls of the counter).
Dr. Kornberg’s key contributions to nucleic acid research: Demonstration of the antiparallel orientation of complementary strands 10/16/2006 Lecture 9: 1 Detection by autoradiography Silver bromide (AgBr), a soft, pale-yellow, insoluble salt well known (along with other silver halides) for its unusual sensitivity to light. This property has allowed silver halides to become the basis of modern photographic materials. The photographic emulsion is usually 10 to 30 µm thick, and is composed of silver halide grains dispersed within gelatin. The grains are 1 µm or greater in diameter; large grains facilitate greater sensitivity, small grains enable finer resolution. The grains consist of silver, bromine, and iodine ions arranged in a crystal lattice (see Fig. 2). Sulfur-containing compounds are often added in order to form specks silver sulfide, which increase photosensitivity.
Dr. Kornberg’s key contributions to nucleic acid research: Demonstration of the antiparallel orientation of complementary strands 10/16/2006 Lecture 9: 3 Detection by autoradiography Chemical properties of photographic film The film base is usually plastic such as tri-acetate or polyester which is coated with a light sensitive emulsion. The emulsion consists of gelatin containing light sensitive silver halide crystals such as silver bromide and silver chloride. In practice the film will consist of many other layers. Photographic emulsion is not a true emulsion, it is a dispersion of small solid particles in a liquid medium which is then allowed to cool and set. The light sensitive crystals are prepared by the combination of silver-Ag- and a halogen. Due to the very low solubility of silver halides mixing aqueous solutions of silver ions and halide ions will result in the precipitation of silver halide crystals. e.g. silver nitrate (AgNO3) + potassium bromide -----> silver bromide (AgBr) + potassium nitrate (KNO3) Or Ag+ (silver ion in solution) + Br- (bromide ion in solution) --------> Ag+Br- (silver bromide crystal) Silver bromide is a lattice crystal containing millions of pairs of ions.
Dr. Kornberg’s key contributions to nucleic acid research: Demonstration of the antiparallel orientation of complementary strands 10/16/2006 Lecture 9: 4 Detection by autoradiography Formation of the Latent Image Step 1: Light Activation It is not fully understood exactly what happens during exposure but the energy released when a photon of light strikes a silver halide crystal frees an electron from the bromide ion. The former bromide ion is released from the crystal as bromine and is absorbed by the gelatin. Step 2: Movement of electrons The free electron moves through the crystal to a 'sensitivity speck' caused by imperfections in the crystal structure or created during the sensitizing process during manufacture. Step 3: Deposition of Silver Ions This now negatively charged speck attracts positive silver ions which are neutralized to form silver atoms. If enough silver atoms form at a single point then a latent image is created. The latent image is not visible, even under a microscope so the only way to tell if it is present is to chemically develop the film to reveal the image.
Dr. Kornberg’s key contributions to nucleic acid research: Demonstration of the antiparallel orientation of complementary strands 10/16/2006 Lecture 9: 5 Detection by autoradiography Development of the latent image Development: During development the developing agent supplies electrons to the latent image thus attracting and neutralizing silver ions to produce metallic silver which will eventually form a visible image. The latent image acts as a catalyst encouraging development to take place faster in exposed areas. Development takes place in both exposed and unexposed areas of the film just at different rates. Developing agents: Metol, Phenidone, Hydroquinone. Stop: When the predetermined development time has been reached the film is moved from the developer to a 'stop bath' which neutralizes the developer and prevents any further development of the image from taking place. Developers work most effectively in an alkaline environment which is why an acid stop bath is used. Stop bath: 1% solution acetic acid. Fixing: After development the emulsion still contains unexposed and undeveloped silver halides. The film will look cloudy or milky and given exposure to light the remaining silver halides will be reduced to silver. The fixer, commonly sodium thiosulphate, converts the unexposed silver halide to soluble salts which can be washed out of the emulsion. Washing: The processed film is washed thoroughly to remove any chemical residue before being dried.
Dr. Kornberg’s key contributions to nucleic acid research: Discovery of poly(P) 10/16/2006 Lecture 9: 6 We (Dr. Arthur Kornberg and his associates) have switched the focus of our research from DNA replication to an entirely new subject: inorganic polyphosphate (poly P). Poly P is a linear polymer of many tens or hundreds of orthophosphate (Pi) residues linked by high-energy, phosphoanhydride bonds. Likely a prominent precursor in prebiotic evolution, poly P is now found in volcanic condensates, deep-oceanic steam vents and in every living thing-bacteria, fungi, protozoa, plants and mammals. Ignored in textbooks and dismissed as a "molecular fossil," our mission is to bring this molecule back to life and demonstrate that poly P is truly a "molecule for many reasons." Our approach is to discover the enzymes for the synthesis and utilization of poly P in bacteria, yeast and animal cells. These enzymes will reveal novel mechanisms and insights and when purified will open the route of reverse genetics: the peptide sequence leads to the gene and thereby the means to knock it out and overexpress it. By manipulating expression of the gene and the cellular levels of its product, phenotypes are created which provide clues to metabolic functions. Most immediate and decisive, the enzymes provide unique and invaluable reagents for analytic and preparative work. Among the several current directions are: DNA entry into cells: The mechanism whereby the inclusion of poly P in a membrane complex enables a cell to become competent to take up DNA and then genetically transformed. Survival in the stationary phase: The basis for poly P regulation of cellular responses to stresses and adjustments for survival in the stationary phase of culture growth and development. In view of the universality and complexity of basic biochemical mechanisms, it would be surprising if some of the variety of poly P functions already observed in microorganisms did not apply to aspects of human growth and development, to aging and to the aberrations of disease. Regulation of development: Developmental changes in microorganisms-fruiting body and spore formation in Myxobacteria (e.g., M. xanthus), sporulation in bacteria (e.g., Bacillus) and fungi, and heterocyst formation in cyanobacteria (e.g., Anabaena)-occur in response to starvation of one or another nutrient. In view of the involvement of poly P in the stationary stage of E. coli, poly P may well participate in other instances of cellular adjustments to deprivation.
Dr. Kornberg’s key contributions to nucleic acid research: 10/16/2006 Lecture 9: 7 He (Dr. Authur Kornberg) was married in 1943 to Sylvy Ruth Levy, who died in 1986. He has three sons and eight grandchildren. Roger is a Professor of Structural Biology at Stanford; Thomas is a Professor of Biochemistry and Biophysics at the University of California in San Francisco; Kenneth is an architect and founder of Kornberg Associates in Menlo Park and Delmar, California, specializing in laboratory design. http://kornberg.stanford.edu/
Dr. Kornberg’s key contributions to nucleic acid research: 10/9/2006 Lecture 9: 8 XI. Good offspring
1. Hexokinase catalyzes: glucose + ATP glucose-6-phosphate + ADP 2. Phosphoglucose Isomerase catalyzes: glucose-6-phosphate (aldose) fructose-6-phosphate (ketose) Fermentation 10/9/2006 Lecture 9: 10
Fermentation 10/9/2006 Lecture 9: 9 3. Phosphofructokinase catalyzes: fructose-6-phosphate + ATP fructose-1,6-bisphosphate + ADP 4. Aldolase catalyzes: fructose-1,6-bisphosphate dihydroxyacetone phosphate + glyceraldehyde-3-phosphate.
Fermentation 10/9/2006 Lecture 9: 11 5. Triose Phosphate Isomerase (TIM) catalyzes:: dihydroxyacetone phosphate (ketose) glyceraldehyde-3-phosphate (aldose) 6. Glyceraldehyde-3-phosphate Dehydrogenase (GAPDH) catalyzes: glyceraldehyde-3-phosphate + NAD+ + Pi 1,3,bisphosphoglycerate + NADH + H+
Fermentation 10/9/2006 Lecture 9: 12 7. Phosphoglycerate Kinase catalyzes: 1,3-bisphosphoglycerate + ADP 3-phosphoglycerate + ATP 8. Phosphoglycerate Mutase catalyzes: 3-phosphoglycerate 2-phosphoglycerate
Fermentation 10/9/2006 Lecture 9: 13 9. Enolase catalyzes: 2-phosphoglycerate phosphoenolpyruvate + H2O 10. Pyruvate Kinase catalyzes: phosphoenolpyruvate + ADP pyruvate + ATP
Fermentation 10/9/2006 Lecture 9: 14 Balance sheet for high energy bonds of ATP: 2 ATP consumed 4 ATP produced (2 from each of two 3C fragments from glucose) Net production of 2 ~P bonds of ATP per glucose. Glycolysis Pathway (omitting H+): glucose + 2 NAD+ + 2 ADP + 2 Pi à 2 pyruvate + 2 NADH + 2 ATP
Fermentation 10/9/2006 Lecture 9: 15 Because glycolysis produces two pyruvate molecules from one glucose, each glucose is processes through the kreb cycle twice. For each molecule of glucose, six NADH2+, two FADH2, and two ATP. C6H12O6 + 6O2 6CO2 + 6H2O + energy (ATP)
Fermentation 10/9/2006 Lecture 9: 16 Fermentation Anaerobic organisms lack a respiratory chain. They must reoxidize NADH produced in Glycolysis through some other reaction, because NAD+ is needed for the Glyceraldehyde-3-phosphate Dehydrogenase reaction (see above). Usually NADH is reoxidized as pyruvate is converted to a more reduced compound. The complete pathway, including Glycolysis and the re-oxidation of NADH, is called fermentation. Skeletal muscles ferment glucose to lactate during exercise. Lactate released to the blood may be taken up by other tissues, or by skeletal muscle after exercise, and converted via Lactate Dehydrogenase back to pyruvate, which may be oxidized in Krebs Cycle or (in liver) converted to back to glucose via gluconeogenesis. Lactate serves as a fuel source for cardiac muscle as well as brain neurons. Astrocytes, which surround and protect neurons in the brain, ferment glucose to lactate and release it. Lactate taken up by adjacent neurons is converted to pyruvate that is oxidized via Krebs Cycle.
Glycogen is a polymer of glucose residues linked mainly by α(1 4) glycosidic linkages. There are α(1 6) linkages at branch points. The chains and branches are longer than shown. Glucose is stored as glycogen predominantly in liver and muscle cells. Fermentation 10/9/2006 Lecture 9: 17 1) Glycogen phosphorylase 2) glucan transferase
Fermentation 10/9/2006 Lecture 9: 18 Pathways involved in the regulation of glycogen phosphorylase. See the text for details of the regulatory mechanisms. PKA is cAMP-dependent protein kinase. PPI-1 is phosphoprotein phosphatase-1 inhibitor. Whether a factor has positive (+ve) or negative (-ve) effects on any enzyme is indicated. Briefly, phosphorylase b is phosphorylated, and rendered highly active, by phosphorylase kinase. Phosphorylase kinase is itself phosphorylated, leading to increased activity, by PKA (itself activated through receptor-mediated mechanisms). PKA also phosphorylates PPI-1 leading to an inhibition of phosphate removal allowing the activated enzymes to remain so longer. Calcium ions can activate phosphorylase kinase even in the absence of the enzyme being phosphorylated. This allows neuromuscular stimulation by acetylcholine to lead to increased glycogenolysis in the absence of receptor stimulation.
Fermentation 10/9/2006 Lecture 9: 19 invertase
net/nfs0/vol1/production/w3no ------TIKVGINGFGRIGRMVFQALCDDGLLG----------------- 27 G3PG_TRYBB ------TIKVGINGFGRIGRMVFQALCDDGLLG----------------- 27 G3PG_TRYCR -----MPIKVGINGFGRIGRMVFQALCEDGLLG----------------- 28 G3PG_LEIME -----APIKVGINGFGRIGRMVFQAICDQGLIG----------------- 28 G3P_TREPA -------MRVAINGFGRIGRLVLQAMAEQKLLG----------------- 26 G3PC_CRAPL ----MAKVKIGINGFGRIGRLVARVALVRDD------------------- 27 G3PC_PETCR -----MKMKIGINGFGRIGRLVARVALMSDD------------------- 26 G3P2_SCHPO ----MAIPKVGINGFGRIGRIVLRNAILTGK------------------- 27 G3PX_HORVU ----MGKIKIGINGFGRIGRLVARVALQSDD------------------- 27 G3P_ATRNU ----MAKVKIGINGFGRIGRLVARVILQSDD------------------- 27 G3P1_ANASQ ---MLAKLKVGINGFGRIGRLVLRAGINNPN------------------- 28 G3P_EMENI -----MAPKVGINGFGRIGRIVFRNAIEAGT------------------- 26 G3PD_MAIZE ----MGKIKIGINGFGRIGRLVARVALQSED------------------- 27 G3PC_MAIZE ----MGKIKIGINGFGRIGRLVARVALQSED------------------- 27 G3PC_GINBI -MSSTGKIKIGINGFGRIGRLVARVALLRDD------------------- 30 G3P1_SCHPO ----MAIPKVGINGFGRIGRIVLRNALVAKT------------------- 27 G3PC_DIACA ----MAPIKIGINGFGRIGRLVARVILQRED------------------- 27 G3P_ASPNG -----MAPKVGINGFGRIGRIVFRNAINHGE------------------- 26 G3P_USTMA ----MSQVNIGINGFGRIGRIVFRNSVVHNT------------------- 27 G3PC_SINAL ---ADKKIKIGINGFGRIGRLVARVILQRND------------------- 28 G3PC_PINSY -MGSTGKIKIGINGFGRIGRLVARVALTRDD------------------- 30 G3PC_PHYPA -MVGSAKIKVGINGFGRIGRLVARVALERDD------------------- 30 G3PC_PETHY ----MAKIKIGINGFGRIGRLVARVALQRDD------------------- 27 G3PC_ANTMA ----MAPIKIGINGFGRIGRLVARVALQRDD------------------- 27 G3PC_ORYSA ----MGKIKIGINGFGRIGRLVARVALQSED------------------- 27 G3PC_MESCR ----MAKVKVGINGFGRIGRLVARVILQRDD------------------- 27 G3PE_MAIZE ----MAKIKIGINGFGRIGRLVARVALQSDD------------------- 27 G3PC_MAGLI --MGGKKIKIGINGFGRIGRLVARVALQRDD------------------- 29 G3PC_RANAC ---MTGKIKIGINGFGRIGRLVARVALARDD------------------- 28 G3PC_GRAVE ----MTVPQVGINGFGRIGRLVLRAAIEKDT------------------- 27 G3PC_ARATH --MADKKIRIGINGFGRIGRLVARVVLQRDD------------------- 29 G3PC_PEA ---MGAKIKIGINGFGRIGRLVARVALKRDD------------------- 28 G3P_ZYGRO ------MVNVSVNGFGRIGRLVTRIAISRKD------------------- 25 G3P1_SYNY3 ------MLKIGINGFGRIGRLVARIAMANPQ------------------- 25 G3P_ERYGR ----MAPIKVGINGFGRIGRIVFRNAAQSCE------------------- 27 G3P_COCHE -----MVVKVGINGFGRIGRIVFRNAIEHND------------------- 26 G3PC_TAXBA -MGSTGKIKIGINGFGRIGRLVARVALQRDD------------------- 30 G3P_PODAN -----MTVKVGINGFGRIGRIVFRNAVEHPD------------------- 26 G3P_PHARH -----MAVKVGINGFGRIGRIVLRNAIIHGD------------------- 26 G3P_KLULA ------MVKVAINGFGRIGRLVLRIALQRKA------------------- 25 G3P2_TRIKO -----APIKVGINGFGRIGRIVFRNAVEHPD------------------- 26 G3P2_AGABI ------MVKVGINGFGRIGRIVLRNALQFQD------------------- 25 G3P1_DROME ----MS--KIGINGFGRIGRLVLRAAIDKG-------------------- 24 G3P_PALVE -------SKIGINGFGRIGRLVLRAALEMG-------------------- 23 G3PC_CHLRE ---MSGKIKIGINGFGRIGRLVMRATMLRPD------------------- 28 G3P_PHYIN -------MNVAINGFGRIGRLVLRASAKNPL------------------- 24 G3P_HOMAM -------SKIGINGFGRIGRLVLRAALSCG-------------------- 23 G3P_SCHCO -----MAVKVGINGFGRIGRIVLRNALQLGN------------------- 26 G3PC_TOBAC ---------------GRIGRLVARVALQRDD------------------- 16 G3P_CURLU -----MVVKVGINGFGRIGRIVFRNAIEHND------------------- 26 G3PC_LEIME -------VKVGINGFGRIGRVVFRAAQMRPD------------------- 24 G3P2_DROME ----MS--KIGINGFGRIGRLVLRAAIDKG-------------------- 24 G3P_TRIHA -----MSIKVGINGFGRIGRILLSNALEKPE------------------- 26 G3P_CANAL -----MAIKIGINGFGRIGRLVLRVALGRKD------------------- 26 G3P_BRUMA ----MSKPKVGINGFGRIGRLVLRAAVEKDT------------------- 27 G3P_PICAN -----MTANVGINGFGRIGRLVLRIALSRDD------------------- 26 G3P_LYOSH ------MVNVGINGFGRIGRIVFRNALLNPK------------------- 25 G3PC_CHOCR ----MTAPKVGINGFGRIGRLVLRAAIEKGT------------------- 27 G3PT_HUMAN MVSVARELTVGINGFGRIGRLVLRACMEKG-------------------- 98 G3P_DROHY ----MS--KIGINGFGRIGRLVLRAAVDKG-------------------- 24 G3P_PHACH -----MPVKAGINGFGRIGRIVLRNALLHGD------------------- 26 G3P_ONCVO ----MSKPKIGINGFGRIGRLVLRAAVEKDT------------------- 27 G3P_PICPA -----MAITVGINGFGRIGRLVLRVALSRAD------------------- 26 G3P_COTJA -------VKVGVNGFGRIGRLVTRAAVLSG-------------------- 23 G3P_CRIGR -------VKVGVNGFGRIGRLVTRAAFTSG-------------------- 23 G3P_CHICK -------VKVGVNGFGRIGRLVTRAAVLSG-------------------- 23 G3P2_HUMAN -----GKVKVGVNGFGRIGRLVTRAAFNSG-------------------- 25 G3P_CRYPA -----MVVKVGINGFGRIGRIVFRNAHEHSD------------------- 26 G3P_PIG -------VKVGVNGFGRIGRLVTRAAFNSG-------------------- 23 G3P_CLAPU -----MAVKVGINGFGRIGRIVFRNAVEHPE------------------- 26 G3P_XENLA -------VKVGINGFGCIGRLVTRAAFDSG-------------------- 23 G3P1_YEAST -------IRIAINGFGRIGRLVLRLALQRKD------------------- 24 G3P_SCHMA ----MSRAKVGINGFGRIGRLVLRAAFLKNT------------------- 27 G3P3_CAEEL ----MTKPSVGINGFGRIGRLVLRAAVEKDS------------------- 27 G3P3_CAEBR ----MSKPSVGINGFGRIGRLVLRAAVEKDS------------------- 27 G3P_RABIT -------VKVGVNGFGRIGRLVTRAAFNSG-------------------- 23 G3P_NEUCR -----MVVKVGINGFGRIGRIVFRNAIEHDD------------------- 26 G3P_ONCMY ----MSDLCVGINGFGRIGRLVLRACLQKG-------------------- 26 G3P_COLGL ----MAPIKVGINGFGRIGRIVFRNAIEHPE------------------- 27 G3PC_TRYBB ------VIRVGINGFGRIGRVVFRAAQRRND------------------- 25 G3P_MOUSE -------VKVGVNGFGRIGRLVTRAAICSG-------------------- 23 G3P2_YEAST -------VRVAINGFGRIGRLVMRIALQRKN------------------- 24 G3P1_JACOR ------MVKVGVNGFGRIGRLVTRAAFNSGKVDIVAINDPFIDLNYMVYM 44 G3P2_CAEEL ----MPKPNVGINGFGRIGRLVLRAAVEKDS------------------- 27 G3P1_CAEEL ----MSKANVGINGFGRIGRLVLRAAVEKDT------------------- 27 G3P_COLLN ----MAPIKVGINGFGRIGRIVFRNAVEHPD------------------- 27 G3P4_CAEEL ----MSKANVGINGFGRIGRLVLRAAVEKDT------------------- 27 G3PT_MOUSE LQKPARELTVGINGFGRIGRLVLRVCMEKG-------------------- 130 G3P2_CAEBR ----MSKPTVGINGFGRIGRLVLRAAVEKDS------------------- 27 G3P1_ECOLI ------TIKVGINGFGRIGRIVFRAAQKRSD------------------- 25 G3P_COLLI -------VKVGVNGFGRIGRLVTRAAILSA-------------------- 23 G3P1_HUMAN -----GKVKVGVDGFGRIGRLVTRAAFNSG-------------------- 25 G3P3_YEAST -------VRVAINGFGRIGRLVMRIALSRPN------------------- 24 G3P_RAT -------VKVGVNGFGRIGRLVTRAAFSCD-------------------- 23 G3P1_GIALA ---MP--IRLGINGFGRIGRMALRASLNIDG------------------- 26 G3P1_TRIKO ------VPKVGINGFGRIGRVVLRNALETGA------------------- 25 G3P2_KLUMA ------MVRIAINGFGRIGRLVLRIALSRKN------------------- 25 G3P_BOVIN -------VKVGVNGFGRIGRLVTRAAFNSG-------------------- 23 G3P_MONAN ----MVVPKVGINGFGRIGRIVFRNAIEHEG------------------- 27 G3PC_HORVU -------------------------------------------------- G3P_SHEEP -----------------IGRLVTRAAFNTG-------------------- 13 Fermentation 10/9/2006 Lecture 9: 20 Evolution of GAPDH