Download

1 / 24
260 likes | 532 Vues
Molecular Markers for Failure of Sulfadoxine-Pyrimethamine Treatment of Plasmodium falciparum and P. vivax Malaria. Zimmerman Lab Melanie V. Prarat Case Western Reserve University 14 July 2004. Mendel and Peas.

E N D
Molecular Markers for Failure of Sulfadoxine-Pyrimethamine Treatment of Plasmodium falciparumand P. vivax Malaria Zimmerman Lab Melanie V. Prarat Case Western Reserve University 14 July 2004
Mendel and Peas • By observing several generations of plants, Mendel discovered that certain traits show up in offspring plants without any blending of parent characteristics • Three important conclusions: • Inheritance of each trait is determined by genes that are passed on to descendents unchanged • An individual inherits one gene for each trait from each parent • A trait may not show up in an individual but can be passed on to the next generation
Genetics • Plasmodium parasites undergo a normal Mendelian pattern of inheritance of genetically determined characters • Blood forms are haploid • Resistance to drugs is due to gene mutations • Recombination between genes determining characters, like drug sensitivity, occurs readily after cross-fertilization between clones • Multi-strain infections provide the principal mechanism for generating parasites with different recombinations of genotypes
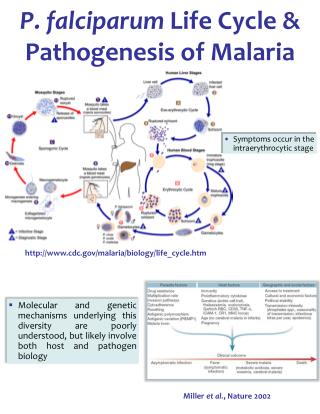
Back to the Basics • Review: • Plasmodium has a complex life cycle • asexual multiplication and gametocyte development occur in the vertebrate host • fertilization between gametes takes place in the mosquito vector
Genetic recombination occurs at a high frequency, resulting in the production of parasites with new genotypes 3D7 x HB3 ↓ >90% recombinants Study showed that two genetically distinct clones of falciparum can undergo mating during mosquito transmission Pyrimethamine was given as the anti-malarial treatment → The recombinant progeny with the HB3 enzyme and antigen characteristics survived What Walliker has to say…
A Brief History of Fansidar® • Sulphadoxine-pyrimethamine (SP) was introduced in 1967 as a synergistic anti-malarial drug, with reports of resistance occurring within a few years • SP (Fansidar®) is used as a second-line drug in areas where chloroquine is still used • In areas that are chloroquine resistant, SP has become the first-line antimalarial • There are many hyperendemic and holoendemic areas that are now SP resistant
Classification of Drug Sensitivity • Used to assess antimalarial susceptibility • High percentages of RII/RIII responses to SP in endemic areas have been documented as early as 1994 in Tanzania • Also found in SE Asia, southern China, & the Amazon Basin • High frequencies of RI/RII responses have been reported on the Pacific coast of South America, southern Asia east of Iran, and throughout sub-Saharan Africa and Oceania • Resistance in Thailand started in the early 1970s • High-level resistance to P.vivax has also been documented in Indonesia and New Guinea, and more recently, in Central America
Another Walliker study… • Showed through a genetic cross that pyrimethamine drug resistance is linked to a point mutation to Asn-108 in the DHFR-TS gene • RFLP analysis showed that there was a HinfI fragment of 4.3kb in the 3D7 parent, and a 4.2kb fragment in the HB3 parents • All pyrimethamine-resistant progeny inherited the HB3 RFLP, and all sensitive progeny inherited that 3D7 RFLP • Identified two other point mutations that confer higher levels of resistance
Dihydrofolate Reductase • DHFR is a small enzyme that plays an essential role in DNA synthesis • Catalyzes the conversion of dihydrofolate to tetrahydrofolate, a cofactor required for the biosynthesis of thymidylate, pyrimidine nucleotides, methionine, and glycine DHFR-TS
Pyrimethamine • Pyrimethamine is one of many antifolates used as an antimalarial drug • Analog of dihydrofolate – similar to one of the rings of DHFR • Inhibits one step of the folic acid synthetic pathway, preventing synthesis of pyrimidines • Leads to inhibition of DNA synthesis
Mechanisms of Resistance • Many theories • Amplification of the DHFR-TS gene • Overexpression of DHFR • Modification of transport of various substrates involved in folate biosynthesis • Drug efflux • P. vivax – inherent resistance to antifolates due to conformation of the active site cavity • Point mutations resulting in the substitution of amino acid residues that form the active site of the DHFR-TS enzyme lead to decreased affinity between antifolates and the enzyme
DHFR Polymorphisms in P. falciparum • Mutations in DHFR confer resistance to pyrimethamine and the other DHFR inhibitors • Ser→Asn 108 causes moderate pyrimethamine resistance in vitro, with higher level resistance resulting from Asn→Ile 51 and/or Cys→Arg 59, and the addition of Ile→Leu 164 causes the highest level of resistance. • DHFR Ser→Asn 108 is used as a marker for SP resistance because it is selected for by SP treatment
P. falciparum Polymorphisms • The substitution of asparagine or threonine at residue 108 reduces the drug’s affinity without affecting the enzyme’s operation on its natural substrate • Multiple mutations diminish the efficacy of the enzyme on the dihydrofolate • Suggests that multiple mutations are unfavorable to the parasites in the absence of drug pressure
DHFR Polymorphisms in P. vivax • I13L • Wild Type 5’ MEDLSDVFDIYAICACCKVA 3’ • Mutant 5’ MEDLSDVFDIYALCACCKVA 3’ • F57L, S58R • Wild Type 5’ SVDMKYFSSVTTYVDESKYE 3’ • Mutant 5’ SVDMKYFRSVTTYVDESKYE 3’ (Burma) • Mutant 5’ SVDMKYLRSVTTYVDESKYE 3’ (Thailand) • S117N/T • Wild Type 5’ VVMGRSSWESIPKQYKPLPN 3’ • Mutant 5’ VVMGRSNWESIPKQYKPLPN 3’ (Burma) • Mutant 5’ VVMGRSTWESIPKQYKPLPN 3’ (Thailand)
P. vivax Polymorphisms • Mutations in P. vivax dhfr codons 58 and 117 are considered equivalent to P. falciparumdhfr residues 59 and 108, respectively • Mutations at 117 in vivax and 108 in falciparum arise first under drug pressure • Highly mutated genes carry the Ser→Thr 117 mutation • 5’ AGC AGC TGG 3’ (WT) • 5’ AGC AAC TGG 3’ • 5’ AGC ACC TGG 3’ ← highly mutated
P. falciparum vs. P. vivax P. vivax 5’ TTC AGG TCG 3’ P. falciparum 5’ TTT CGT GCA 3’ • P. vivax S58R • P. falciparum C59R P. vivax 5’ AGC ACC TGG 3’ P. vivax 5’ AGC AAC TGG 3’ P. falciparum 5’ ACA AAC TGG 3’ P. falciparum 5’ ACA ACC TGG 3’ • P. vivax S117T/N • P. falciparum S108T/N
Interpretation and Results • The positive controls, HB3 and K1, verified that the BioPlex results were correct, even if the numbers are lower than the average output values • HB3 – 51A, 59T, 108A, 164A • K1 – 51A, 59C, 108A, 164A • All the field samples had higher levels of resistance to sulfadoxine-pyrimethamine than wild-type strains • Since the blood forms of malaria parasites are haploid, it can be presumed that the samples with more than one positive result from the BioPlex machine have more than one parasite strain present • Technique can be used to genotype samples quickly and efficiently → monitor drug resistance in different geographical locales
PNG • According to Casey et al. … • Prior to his 2004 study, chloroquine had been the standard treatment for uncomplicated malaria, and Fansidar in combination with quinine was the second-line treatment • During the course of the study, the PNG Department of Health revised the standard treatment recommendations to chloroquine + SP and SP + artemisinin derivatives • No polymorphisms were observed at codons 16, 51, or 164 of dhfr at either of the two study sites (Madang and Maprik) • There were higher rates of mutations at residues 59 and 108 in Madang, where people have a higher exposure rate to SP • 108 mutations also strongly correlated with prior exposure to primaquine • Once SP becomes ineffective, all that will be left are the artemisinin derivatives
Roll Back of Antifolate Resistance • Studies show that by implementing insecticide-treated nets, there is an increased frequency of P. falciparum strains carrying the wild-type dhfr allele • ITNs reduce transmission, therby reducing the number of drug-exposed parasites
Future Concerns • Areas with no drug-resistant malaria • Efforts are needed to extend chloroquine efficacy for as long as possible • Areas with no multidrug resistance • Constant surveillance needed for the early detection of development of multidrug resistance • Emerging multidrug-resistant areas • Prevention and rapid detection of new foci development • New regimens and alternative drugs needed • Rational drug use must be reinforced to limit drug pressure • Established multidrug-resistant areas • Efforts needed to limit geographical spread • New combinations may preserve efficacy of current drugs • Development of new drug candidates needed
Alifrangis, M. et al.. Am. J. Trop. Med. Hyg.. 2003;69(3): 238-43. Diourté, Y. et al.. American Journal of Tropical Medicine and Hygiene. 1999;60(3): 475-78. Imwong, M. et al.. Antimicrobial Agents and Chemotherapy. 2003;47:1514-21. Kublin, James G. et al.. The Journal of Infectious Diseases. 2002;185: 380-8. Le Bras, J., & R. Durand. Fundamental & Clinical Pharmacology. 2003;17: 147-53. Nirmalan, N. et al.. Mol. Microbiol. 2002 Oct;46(1):179-90. de Pécoulas, P.E. et al.. Molecular and Biochemical Parasitology. 1998;92: 265-73 de Pécoulas, P.E. et al.. Gene. 1998;211: 177-85. Sibley, C. et al.. Trends in Parasitology. 2001; 17(12): 582-88. Walliker, D. et al.. Science. 1987;236: 1661-66. Peterson, D.S. et al.. Proc. Natl. Acad. Sci. 1988;85: 9114-118. Wang, P. et al. Molecular and Biochemical Parasitology. 1995;71: 115-25. Wongsrichanalai, C. et al.. The Lancet Infectious Diseases. 2002;2: 209-18. Zimmerman, P. Am. J. Trop. Med. Hyg.. 2003;69(3): 236-37. Works Cited