Download
1 / 1
10 likes | 107 Vues
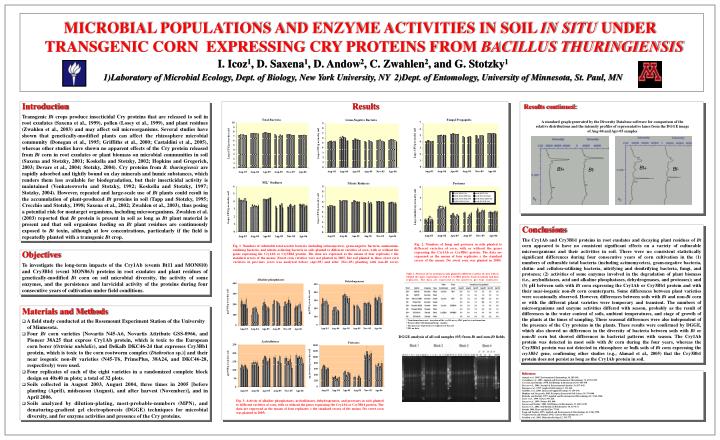
Bt +. Bt -. Bt +. Bt -. DGGE analysis of all soil samples (05) from Bt and non- Bt fields. MICROBIAL POPULATIONS AND ENZYME ACTIVITIES IN SOIL IN SITU UNDER TRANSGENIC CORN EXPRESSING CRY PROTEINS FROM BACILLUS THURINGIENSIS.

E N D
Bt+ Bt- Bt+ Bt- DGGE analysis of all soil samples (05) from Bt and non-Bt fields MICROBIAL POPULATIONS AND ENZYME ACTIVITIES IN SOIL IN SITU UNDER TRANSGENIC CORN EXPRESSING CRY PROTEINS FROM BACILLUS THURINGIENSIS I. Icoz1, D. Saxena1, D. Andow2, C. Zwahlen2, and G. Stotzky1 1)Laboratory of Microbial Ecology, Dept. of Biology, New York University, NY 2)Dept. of Entomology, University of Minnesota, St. Paul, MN Introduction Results Transgenic Bt crops produce insecticidal Cry proteins that are released to soil in root exudates (Saxena et al., 1999), pollen (Losey et al., 1999), and plant residues (Zwahlen et al., 2003) and may affect soil microorganisms. Several studies have shown that genetically-modified plants can affect the rhizosphere microbial community (Donegan et al., 1995; Griffiths et al., 2000; Castaldini et al., 2005), whereas other studies have shown no apparent effects of the Cry protein released from Bt corn in root exudates or plant biomass on microbial communities in soil (Saxena and Stotzky, 2001; Koskella and Stotzky, 2002; Hopkins and Gregorich, 2003; Devare et al., 2004; Stotzky, 2004). Cry proteins from B. thuringiensis are rapidly adsorbed and tightly bound on clay minerals and humic substances, which renders them less available for biodegradation, but their insecticidal activity is maintained (Venkateswerlu and Stotzky, 1992; Koskella and Stotzky, 1997; Stotzky, 2004). However, repeated and large-scale use of Bt plants could result in the accumulation of plant-produced Bt proteins in soil (Tapp and Stotzky, 1995; Crecchio and Stotzky, 1998; Saxena et al., 2002; Zwahlen et al., 2003), thus posing a potential risk for nontarget organisms, including microorganisms. Zwahlen et al. (2003) reported that Bt protein is present in soil as long as Bt plant material is present and that soil organisms feeding on Bt plant residues are continuously exposed to Bt toxin, although at low concentrations, particularly if the field is repeatedly planted with a transgenic Bt crop. Results continued: A standard graph generated by the Diversity Database software for comparison of the relative distributions and the intensity profiles of representative lanes from the DGGE image of Aug-04 and Apr-05 samples Conclusions The Cry1Ab and Cry3Bb1 proteins in root exudates and decaying plant residues of Bt corn appeared to have no consistent significant effects on a variety of culturable microorganisms and their activities in soil. There were no consistent statistically significant differences during four consecutive years of corn cultivation in the (1) numbers of culturable total bacteria (including actinomycetes), gram-negative bacteria, chitin- and cellulose-utilizing bacteria, nitrifying and denitrifying bacteria, fungi, and protozoa; (2) activities of some enzymes involved in the degradation of plant biomass (i.e., arylsulfatases, acid and alkaline phosphatases, dehydrogenases, and proteases); and (3) pH between soils with Bt corn expressing the Cry1Ab or Cry3Bb1 protein and with their near-isogenic non-Bt corn counterparts. Some differences between plant varieties were occasionally observed. However, differences between soils with Bt and non-Bt corn or with the different plant varieties were temporary and transient. The numbers of microorganisms and enzyme activities differed with season, probably as the result of differences in the water content of soils, ambient temperatures, and stage of growth of the plants at the times of sampling. These seasonal differences were also independent of the presence of the Cry proteins in the plants. These results were confirmed by DGGE, which also showed no differences in the diversity of bacteria between soils with Bt or non-Bt corn but showed differences in bacterial patterns with season. The Cry1Ab protein was detected in most soils with Bt corn during the four years, whereas the Cry3Bb1 protein was not detected in rhizosphere or bulk soils of Bt corn expressing the cry3Bb1 gene, confirming other studies (e.g., Ahmad et al., 2005) that the Cry3Bb1 protein does not persist as long as the Cry1Ab protein in soil. Fig. 2. Numbers of fungi and protozoa in soils planted to different varieties of corn, with or without the genes expressing the Cry1Ab or Cry3Bb1 protein. The data are expressed as the means of four replicates ± the standard errors of the means (No sweet corn was planted in 2005). Fig. 1. Numbers of culturable total aerobic bacteria (including actinomycetes), gram-negative bacteria, ammonium-oxidizing bacteria, and nitrate-reducing bacteria in soils planted to different varieties of corn, with or without the genes expressing the Cry1Ab or Cry3Bb1 protein. The data are expressed as the means of four replicates ± the standard errors of the means (Sweet corn varieties were not planted in 2005, but soil planted to these sweet corn varieties in previous years was analyzed before (Apr-05) and after (Nov-05) planting with non-Bt corn). Objectives To investigate the long-term impacts of the Cry1Ab (events Bt11 and MON810) and Cry3Bb1 (event MON863) proteins in root exudates and plant residues of genetically-modified Bt corn on soil microbial diversity, the activity of some enzymes, and the persistence and larvicidal activity of the proteins during four consecutive years of cultivation under field conditions. Table 1. Presence of Cry proteins in soils planted to different varieties of corn, with or without the genes expressing Cry1Ab or Cry3Bb1 protein. Each treatment had four replicates. The data are expressed as the positives per four replicates. Materials and Methods • A field study conducted at the Rosemount Experiment Station of the University of Minnesota. • Four Bt corn varieties [Novartis N45-A6, Novartis Attribute GSS-0966, and Pioneer 38A25 that express Cry1Ab protein, which is toxic to the European corn borer (Ostrinianubilalis), and DeKalb DKC46-24 that expresses Cry3Bb1 protein, which is toxic to the corn rootworm complex (Diabrotica sp.)] and their near isogenic non-Bt varieties (N45-T6, PrimePlus, 38A24, and DKC46-28, respectively) were used. • Four replicates of each of the eight varieties in a randomized complete block design on 40x40 m plots; a total of 32 plots. • Soils collected in August 2003, August 2004, three times in 2005 [before planting (April), midseason (August), and after harvest (November)], and in April 2006. • Soils analyzed by dilution-plating, most-probable-numbers (MPN), and denaturing-gradient gel electrophoresis (DGGE) techniques for microbial diversity, and for enzyme activities and presence of the Cry proteins. a Transformation event: insertion of the cry1Ab or cry3Bb1 gene by transformation. b Western blot with ImmunoStrips (Agdia). C The presence of protein in 4 replicates of the soil. d ND, no data. References: Ahmad et al., 2005. Environmental Entomology 34, 385-394. Castaldini et al., 2005. Applied and Environmental Microbiology 71, 6719-6729. Crecchio and Stotzky, 1998. Soil Biology & Biochemistry 30, 463-470. Devare et al., 2004. Journal of Environmental Quality 33, 837–843. Donegan et al., 1995. Applied Soil Ecology 2, 111-124. Griffiths et al., 2000. Journal of Applied Ecology 37, 159-170. Hopkins and Gregorich, 2003. European Journal of Soil Science 54, 793-800. Koskella and Stotzky, 1997. Applied and Environmental Microbiology 63, 3561-3568. Losey et al., 1999. Nature399, 214. Saxena et al., 1999. Nature 402, 480. Saxena and Stotzky, 2001. Soil Biology & Biochemisty 33, 1225-1230. Saxena et al., 2002. Soil Biology & Biochemistry 34, 133-137. Stotzky 2004. Plant and Soil 266, 77-89. Tapp and Stotzky, 1995. Applied and Environmental Microbiology 61, 1786-1790. Venkateswerlu and Stotzky, 1992. Current Microbiology 25, 1-9. Zwahlen et al., 2003. Molecular Ecology 12, 765-775. Fig. 3. Activity of alkaline phosphatases, arylsulfatases, dehydrogenases, and proteases in soils planted to different varieties of corn, with or without the genes expressing the Cry1Ab or Cry3Bb1 protein. The data are expressed as the means of four replicates ± the standard errors of the means (No sweet corn was planted in 2005).