Download

1 / 1
10 likes | 89 Vues
W. Coul. VdW. D A s. D A r. D A w. D A tot. rms. w1. 12.1. -1.9. 0.4. -3.0. 5.9. 1.7. 0.1. w2. 8.7. -1.4. 0.4. -3.2. 5.9. -1.4. 0.4. w3. 13.4. -1.1. 0.4. -3.2. 5.9. 3.6. 0.1. UIC. w4. 14.9. -1.1. 0.4. -3.2. 5.9. 4.0. 0.4. 3+4. 22.4. 0.7. 0.8. -6.8.

E N D
W Coul VdW DAs DAr DAw DAtot rms w1 12.1 -1.9 0.4 -3.0 5.9 1.7 0.1 w2 8.7 -1.4 0.4 -3.2 5.9 -1.4 0.4 w3 13.4 -1.1 0.4 -3.2 5.9 3.6 0.1 UIC w4 14.9 -1.1 0.4 -3.2 5.9 4.0 0.4 3+4 22.4 0.7 0.8 -6.8 11.8 5.3 0.6 Water FF Coul VdW DAs DAr DAw DAtot rms W1 Amber99 12.1 -1.9 0.4 -3.0 5.9 1.7 0.1 Amber03 11.9 -1.3 0.4 -3.2 5.9 2.0 0.2 W2 Amber99 8.7 -1.4 0.4 -3.2 5.9 -1.4 0.4 Amber03 8.7 -1.9 0.4 -3.2 5.9 -1.9 0.1 OPLS-AA 9.0 -1.8 0.4 -3.2 5.9 -1.5 0.8 Binding Free Energies of Water Molecules in Protein Active Sites A Case Study Using Pentothenate Synthetase, An Emerging TB target Michael Brunsteiner, Reaz Uddin, Pavel A. Petukhov* (email: pap4@uic.edu) Department of Medicinal Chemistry and Pharmacognosy, College of Pharmacy, University of Illinois at Chicago 833, S. Wood St. Chicago, IL 60612 INTRODUCTION Crystal structures of protein-ligand complexes frequently show the presence of water molecules in binding sites. Replacement of such water molecules through modification of ligand molecules can both increase or decrease the binding affinity of ligands, depending on the free energy cost required to transfer the water from the binding site to the bulk. For the purpose of lead optimization it can therefore be beneficial to know the binding affinities of such water molecules. Here we present results obtained for Pantothenate Synthetase (PS), an emerging target for the treatment of tuberculosis. We performed extensive free energy calculations comparing the binding affinities of conserved water molecules in four different locations in the PS binding site. We investigate a number of possible factors that may influence the accuracy of such calculations and discuss implications of our results for the design of new PS inhibitors. PROTEIN AND WATER PS catalyzes the formation of pantothenate from ATP, D-pantoate and β-alanine in Mycobacterium tuberculosis. Several crystal structures of this protein complexed with the natural substrate or a reaction intermediate suggest the presence of four water molecules that interact with both ligand and protein. (Figure 1 and 2) COMPUTATIONAL DETAILS Preparation of initial structures: Available crystal structures (1N2I, complex PS-reaction intermediate and 2A88, PS apo-protein) were protonated usingpdb2gmx. Protonation of ionizable groups and ASN/GLN orientations were double-checked using dedicated software (NQ-flipper, H++), and by visual inspection. The protein structures were solvated with 9600 water molecules in cubic box. Na and Cl ions were added at c ~ 0.1 M at a ratio that ensured a system with zero net-charge. MD simulations Simulations in the NVT ensemble were performed using the program Gromacs, with a leap-frog integrator, a time step of 2 fs, a Nose-Hoover thermo-state at 300 K. Electrostatic long range interactions were calculated using the PME algorithm. Harmonic position restraints were applied to all backbone atoms more than 10 Å from any ligand or conserved water atom. Unless stated explicitly AMBER99 was used for the protein and GAFF+AM1BCC charges for the ligand. Calculation of free energies Free energies were calculated by thermodynamic integration. Electrostatic and VdW interactions were turned off separately over 11 and 17 windows, resp. Each window extended at least over 400 ps. Unless stated explicitly each result is an average from four independent simulations stared from different initial structures. Separation shifted scaling was used to avoid numerical instabilities and diverging free energies. Harmonic position restraints were applied to the interesting water atom to ensure convergence of free energies when decoupling it from the system. The total free energy difference between a water in the binding site and in the bulk was calculated as: DA =DACoul +DAVdW + DAs +DAr - DAw Where DACoul and DAVdW are the electrostatic and non-polar contributions to the binding free energy, calculated through thermodynamic integration, DAr is the correction term for the harmonic restraint applied to the water, DAs is an energy term to account for the water symmetry, and DAw is the free energy to transfer a water from vacuum to the bulk. RESULTS The calculated numbers (Table 1) confirm for three of the four water molecules that they are more stable at their location in the binding site than they are in the bulk. The values for W3 and W4 correspond to the same initial and final states. The fact that they are within each others error bars confirms the convergence of these calculations. Replacing W3 or W4 (or both) entails a considerably higher free energy cost than W1 or W2. We conclude that replacing W1 and, in particular, W2 through modification of an appropriate ligand will result in better chances of increased binding affinity of this ligand than replacing W3 and W4. ● THE CONSTRAINTS Figure 3 shows that the fluctuations of the water molecules are very close to spherical: we conclude that the correction term that we apply for the harmonic constraint that keeps the water in place while the interactions are turned off is appropriate. We also calculated the free energy cost of constraining the water explicitly. The calculated value of 0.6 kJ/mol is non-negligible, but is still too small to account for the unexpected energies we see for W2. Figure 3: density distributions of the four water oxigens from 2 ns unconstrained MD simulations ● CONVERGENCE To ensure that no artifacts due to inaccurate estimation of the equilibrium position and RMSD occur we use comparatively small force constants for the harmonic potential constraining the water molecules while their interactions with the system are turned off. This requires longer simulation times, raising the question whether our results are converged. Figure 4 shows that for W2 the results are converged within their respective error-bars. Figure 4: Running averages of the VdW and Coulomb contributions (each 4 runs) calculated for W2. Table 1: The total free energy cost of transferring a water from its location in the binding site into the bulk, and its contributions. In the line labeled 3+4 we put the sum of the average from 3 and 4 plus the energies for removing W3 with no W4 present and vice versa. ● THE PROTEIN STRUCTURE We repeated some of our calculations starting from a different crystal structure of PS, the apo-protein (2A88) into which we modeled through alignment the same ligand used before. We find that the free energy of transferring W2 to the bulk is even more negative than found for 1N2H. IS THERE A W2 ? The calculated free energy difference for transferring a water from the location W2 to the bulk is negative, predicting that there is no water in this location. This result disagrees with the available crystal structures for PS and is possibly an artifact. We investigated a number of possible reasons for this unexpected result. Table 2: Comparison of free energies calculated for waters at positions W1 and W2 for two different structures of PS. ● POLARIZBILITY Here we used force fields with static partial charges throughout. To estimate quantitatively the influence of polarization we calculated the average electrostatic field and its fluctuations at the position of the oxygen atom of the four conserved waters. Comparison with the same quantities calculated for bulk waters (Figure 7) shows that W2 embedded in the PS binding site is unlikely to experience larger polarization forces than bulk water. We conclude that not accounting for polarization is unlikely to cause the unexpected free energy of W2. • SUMMARY AND OUTLOOK • Calculated relative free energies indicate that replacement of a water in position W3 or W4 would entail a relatively large energetic penalty. In the design of PS inhibitors via, e.g., docking these waters should, thus, remain in place. • According to our results a conserved water molecule in position W2, as suggested by a number of x-tal structures, would be unstable. We investigated a range of possible reasons to find that this result is unlikely to be an artifact. We expect the density observed at the location W2 to stem from an buffer ion rather than from water, and currently investigate the implications of this finding for the design of new PS inhibitors. Figure 1: Seven x-tal structures aligned reveal four clusters of conserved water. Figure 5: Electrostatic field (average+fluctuations) on the position of the water oxygens (crystal vs bulk) ● THE FORCE FIELD We repeated some of the calculations with different force fields. The results are given in Table 2. We see some variations, but the absolute free energies are generally within each others error-bars, and the qualitative conclusion does not depend on the force field. ACKNOWLEDGEMENTS This research was supported by the National Institute of Health/National Institute of Allergy and Infectious Diseases grant R21 AI070997 and Institute for Tuberculosis Research at University of Illinois at Chicago. We also thank Higher Education Commission of Pakistan for providing financial support to Reaz Uddin; and David Mobley, Eric Sorin, and John Codera for providing the implementation of Amber and GAFF for Gromacs. Figure 2: Two views of the PS binding site, containing a natural substrate/AMP reaction-intermediate. The conserved waters considered here are colored green and labeled. H-bonds are indicated by blue lines. W3 and W4 are directly next to each other and interacting through an H-bond. Removing only one of W3 or W4 is likely to de-stabilize the other. Thus we assume that only a concerted removal of both waters can decrease the total free energy of the system, and, thus, increase the binding affinity of the ligand. Table 3: The electrostatic field at the position of the four conserved water molecules compared to a bulk water











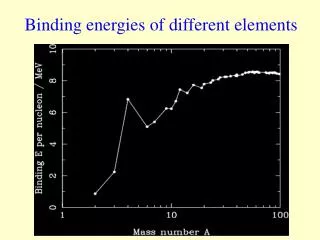
![Binding energies in DATA [MeV]](https://cdn2.slideserve.com/3921996/slide1-dt.jpg)