Download

1 / 27
280 likes | 450 Vues
1. IMA Sept 15th-21st 2007. Nucleosome dynamics probed by torsional manipulation of single chromatin fibers Prunell et al. Force. (600 bp). (500 bp). (2x18 5S 208- or 190-bp). (600 bp). (500 bp). 2. Scheme of the magnetic tweezers set-up. 3.

E N D
1 IMA Sept 15th-21st 2007 Nucleosome dynamics probed by torsional manipulation of single chromatin fibersPrunell et al.
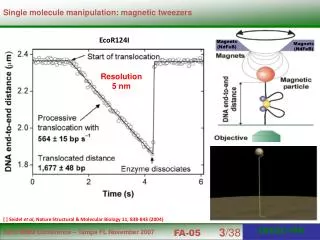
Force (600 bp) (500 bp) (2x18 5S 208- or 190-bp) (600 bp) (500 bp) 2 Scheme of the magnetic tweezers set-up
3 EM of typical 2x18 5S 190 bp chromatin fibers with linkers in TE Clusters of close-packed nucleosomes ~1100-bp nucleosome-free DNA spacers and stickers
4 Part 1A fiber of three-state nucleosomes
5 Force-vs.-extension of a 5S 2x18 208-bp chromatin fiber in TE Unfolding regime Entropic regime Chromatin Naked DNA Smooth curves by the worm-like rope elasticity model (Bouchiat et al. 1998, Phys. Rev. Lett. 80, 1556-1559): Chromatin stretching modulus ~8 pN
6 Extension-vs.-rotation of a 5S 2x18 208-bp chromatin fiber and its corresponding naked DNA in TE Elastic regime Plectonemic regime Strand separation Lengths (µm) Chromatin Naked DNA
7 Fiber parameters before and after partial nucleosome depletion NAP-1 + 50 mM NaCl f = 7.7 pN f = 7.7 pN f = 0.7 pN 14 steps of 24±2 nm = 14 nucleosomes removed Topological shifts ∆(length)/topological shift ~1.35 µm/24 turns = ~55 nm/turn or ~160 bp/turn ~700 nm/13 turns or 54 nm /turn ∆(Topological shift)/∆(nucleosome) ~ -11/14 = -0.8±0.1
Slope ~ 55 nm/turn 8 Regular and irregular fibers 6 Regular fibers contain mostly regularly spaced nucleosomes and are on the straight line Irregular fibers contain a significant number of close-packed nucleosomes of smaller topological deformation
2 9 Fibers are extremely flexible in torsion compared to DNA of the same length 2 Renormalized naked DNA Smooth curve by the worm-like rope model: Bending persistence length ~28 nm Torsional persistence length ~5 nm against 80 nm for DNA
2 7 2 6 10 Irregular fibers are less torsionally flexible 7 6 Slope ~ 55 nm/turn
11 The two-angle molecular model of the canonical (all-negative) 2x18 208-bp fiber between entry/exit DNAs between successive nucleosomes ∆l : rise per nucleosome ∆Lkt : theoretical topological deformation per nucleosome ∆Lkm : experimental deformation for a single nucleosome on a DNA minicircle of the same 5S sequence ∆Lkm(turn) -1.4 Elastic regime Torsional persistence length ~ 35 nm (against 5 nm for the real fiber)
12 Nucleosome three conformational states Recent reviews of the DNA minicircle approach : Prunell A. & Sivolob A. (2004) New Comprehensive Biochemistry, 39, 45-73 Sivolob A. & Prunell A. (2004) Phil. Trans. Roy. Soc. A. 362, 1519 - 1547
13 All-open- and all-positive fibers entry/exit nucleos./nucleos. rise t for theoretical m for minicircle
14 The equilibrium three-state model of the chromatin fiber Elastic regime Fiber 2 Plectonemic regimes The model uses the experimentally-determined number of nucleosomes and, as adjustable parameters, the difference in energy between the states (negative - open = 0.7 kT; positive - open = 2 kT) Slope of the plectonemic regimes: ~ 25 nm/turn against 90 nm/turn for the naked DNA
15 Details of the modelled fiber Torsionally-relaxed fiber (zero torque) negative Steady-state number of nucleosomes (total = 31) open positive Models: Torque (pN.nM/rad) worm-like rope molecular Circles: onsets of plectoneme formation at constant torque The maximal length maximizes the number of open nucleosomes at ~ +5 turns from the relaxed state
16 Part 2The nucleosome chiral transition
Naked DNA 17 The hysteretic torsional response Hysteretic shifts on the positive side ( ) remain constant for a given fiber over many cycleds of rotation, while it varies on the negative side, in particular as a function of the acquisition time The fiber maximal extension is approximately the same inforwardandbackwardcurves Conclusion: The backward curve as a whole tends to shift to larger rotations relative to the forward curve as a consequence of positive turn trapping by the fiber after excursion at high torsion
ntotal nreg 18 Hysteretic shifts are proportional to the number of regularly-spaced nucleosomes (1) (2) Slope = 1.3 ±0.1 turns/reg. nucl. Conclusion: close-packed nucleosomes contribute little to the hysteresis Shift increases at a rate of 1.3 turns per regularly spaced nucleosome. With a topological deformation of the positively-crossed nucleosome in the positive plectonemic regime of ~ -0.4 , the topological deformation of the altered nucleosome form is 1.3 -0.4 = 0.9 ±0.2
19 The altered nucleosome form contains the H2A-H2B dimers 2- removal of H2A-H2B dimers at 3.5 pN with 250 nM yNAP-1 dimers in TE + 50 mM NaCl followed by rinsing in TE and return to 0.2 pN results in a hysteresis-free fiber 3- rescue of initial fiber length and hysteresis upon incubation with 40 nM H2A-H2B dimers chaperoned with 40 nM NAP-1 dimers and rinsing in TE 4- all histones removed (with heparin) Conclusion: The altered nucleosome form must contain H2A-H2B dimers as they are required for the hysteresis
20 An elusive tetrasome fiber Direct reconstitutions with tetramers lead to authentic hairpin-like tetrasomes only at low histone/DNA ratios. Ratios insuring a reasonable particle density along the DNA produce pseudo-nucleosomes made of two stacked (H3-H4)2 tetramers Insets: particles obtained upon reconstitutions on a ~ 200 bp 5S DNA fragment
1 µg/ml heparin + 1 µg/ml core particles as histone acceptors 700 mM NaCl 21 Dimers removed by The true tetrasome fiber Dimers removed Naked DNA An authentic tetrasome fiber (obtained by H2A-H2B removal and not by direct reconstitution) shows : 1) an absence of hysteresis 2) a maximal extension intermediate between the initial chromatin fiber and naked DNA consistent with tetrasome wrapping of less than a turn 3) an absence of topological shift relative to naked DNA. This is consistent with the known ability of single tetrasomes on DNA minicircles to fluctuate between pseudo-mirror-symmetrical left- and right-handed chiral conformations of nearly equal and opposite ∆Lk (-0.7 and + 0.6 (±0.05) on 5S DNA). This in turn implies that the tetrasome fiber at its center of rotation contains an equal steady state number of particles of each chirality
22 The altered nucleosome versus the right-handed tetrasome 1 µg/ml heparin + 1 µg/ml core particles as acceptors 700 mM NaCl 1) The tetrasome fiber and the nucleosome backward fiber have the same center of rotation. 2) Their responses have the same breath. 3) The topological deformation of the altered nucleosome (+0.9) is similar to that of the right-handed tetrasome on the same 5S sequence (+0.6) Thebackward transition process must be similar for the altered nucleosome and the tetrasome. The difference in the forwardresponses may then solely reflect the activation energy of the nucleosome transition linked to the presence of H2A-H2B dimers. Whole nucleosomes also switch their conformation from left-handed to right-handed, and the core of the altered form is the right-handed tetrasome
23 The transition in real time: a test for the metastability of the altered nucleosome A- Fiber length monitored at constant force in the backward curve. The proportions of the different states (inset) is used to estimate the energy parameters of the transition (from the ground state of the open nucleosome) according to standard calculation: - Equilibrium energy difference = 10 ±2 kT - Energy barrier = 30 ±5kT B- Length monitored in the forward curve at constant force. No extension is observed in 30 min, consistent with the >90 % straight nucleosome proportion at steady state (measured from the inset in A at infinite time), and the high energy barrier. A conversion to the altered form is observed only when the equilibrium is displaced toward the altered state by a force >2.3 pN.
24 Salt-dependence of the torsional response and the chiral transition • Fibers in higher salt (25 - 50 mM NaCl) tend to progressively compact from one cycle of torsion to the next. This is due to salt-induced attractive interactions between the nucleosomes, which are mediated by the tails. However, a force of a few pN applied at the center of rotation breaks these interactions and rescues the initial response which is close to that in low salt. • The hysteresis tends to disappear and forward and backward curves tend to merge toward an intermediate curve. Real-time monitoring shows a substantial decrease in the energies: • Equilibrium energy difference = 6 ±2 kT, against 10 ±2 kT in low salt • Energy barrier = 25 ±5 kT, against 30 ±5 kT in low salt. Because the transition rate depends exponentially on the energy barrier, the equilibrium between the two states becomes more dynamics relative to the time-scale of data acquisition, explaining the merging
Nucleosome Reversome positive rotation ∆Lk ~-1 ∆Lk ~+1 25 Putative scenario for the transition Similar fiber extension in forward and backward curves implies a similar length component along the force of canonical and altered nucleosome forms Conclusion : The two forms should wrap the same length of DNA with similar compactions
26 Part 1 Nucleosome access to three conformational states (depending on the crossing status of entry-exit DNAs: negative, null or positive) solves a number of previous topological enigmas of DNA in chromatin. The most famous is the linking number paradox, where two negative superhelical turns around the nucleosome reduce (on average) the linking number by only 1 (instead of 2). The key nucleosomes here are the positively crossed ones, because they are almost topologically neutral and contribute little to the overall linking number reduction. Part 2 Nucleosome chiral transition into reversomes is likely to occur during transcriptional elongation. RNA polymerases exert a positive torque >1.25 kT/rd, i. e. 8 kT/turn, sufficient in principle to trigger the transition (6 kT/turn in 50 mM salt). However, it is the activation energy which specifies the conversion rate, and for this reason it was important to know whether reversomes could be produced at a distance in a time-scale consistent with the polymerase elongation speed. We thus implemented a kinetic model in which a fiber is twisted at constant angular velocity, the torsional constraint being relaxed by the nucleosome- reversome transition in a steady-state manner. Given the speed of RNA polymerase II (~20 nucleotides or 2 turns/sec), the above energy parameters lead to the effective torque involved, ~1.5 kT/rad. This figure, close to the above minimal torque value, suggests that the transition may indeed propagate ahead of a transcribing polymerase. If reversomes indeed form, what could their role be ? H2A-H2B dimers exert an almost absolute block against transcription by RNA polymerase II at physiological ionic strength. This has been observed on short chromatin substrates, when positive supercoiling cannot accumulate due to free rotation of the ends. Reversomes, owing to their open character, have their dimers relatively destabilized, suggesting they could behave as torsion-driven “activated” nucleosomes poised for polymerase traversing. The chiral-switching ability of the tetramer may then be viewed as the lever used by the main RNA polymerase to break the docking of the dimers, via the wave of positive supercoiling it pushes in its front. Physiological relevance of the data
27 Work attributions 1) Jean-Louis Viovy, withAurélien Bancaud and Gaudeline Wagner (Institut Curie-Paris) provided and operated the magnetic tweezers set-up 2) Jean-Marc Victor, withJulien Mozziconacci, Hua Wong and Maria Barbi (LPTMC, Paris) modeled and simulated 3) Eric le Cam and Christophe Lavelle (Institut Gustave Roussy, Villejuif) visualized the fibers 4) Liliane Mouawad (Institut Curie-Orsay)performed the Normal mode analysis of tetrasome chiral transition 5) Andrei Sivolob (Taras Shevchenko National University, Kiev) calculated the energies 6) Ariel Prunell and Natalia Conde e Silva (Institut Jacques Monod, Paris) prepared the chromatin fibers • For more details, see: • Bancaud et al. (2006) Nat. Struct. Mol. Biol. 13, 444-450. • Bancaud et al. (2007) Mol. Cell 27, 135-147. • and for a genealogy of the nucleosome superfamily, see: • Lavelle & Prunell (2007) Cell cycle 6,2113-2119 (Review)