Download

1 / 31

E N D
MODUL/TOPIK 5 FERTILISASIAWAL A. PENDAHULUAN Modul 5 ini menjelaskan mengenai proses fertilisasi dan macam-macam fertilisasi yang dilakukan oleh hewan-hewan Invertebrata maupun Vertebrata. Selain itu dalam mengkaji mekanisme fertilisasi awal ada 2 aktivitas utama, yaitu pengenalan (kontak awal) antara sperma dan sel telur, dan pengaturan pemasukkan sperma ke sel telur, Setelah mempelajari modul ini mahasiswa diharapkan mampu memahami tentang fertilisasi baik macam, mekanisme, dan periodanya pada beberapa hewan dari golongan Invertebrata maupun Vertebrata. Kemudian mahasiswa dapat menjelaskan mekanisme fisiologis pada saat terjadi kontak awal antara spema dan sel telur, serta dapat mendiskusikan masalah kecenderungan bahwa hanya satu sperma saja yang mampu menembus sel telur, dan akan melengkapi diskusi tersebut dengan bahan-bahan yang diperoleh dari internet. B. URAIAN Fertilisasi adalah proses peleburan 2 macam gamet, sehingga terbentuk individu baru dengan sifat genetik yang berasal dari kedua induknya. Prefertilisasi atau fertilisasi awal secara garis besar meliputi 2 aktivitas utama, yaitu (1) Pengenalan (kontak awal) sperma
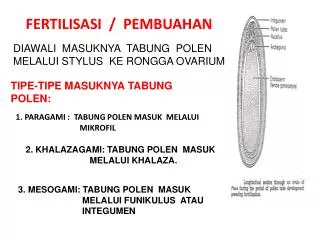
dan sel telur dan (2) Pengaturan pemasukkan sperma ke sel telur (Bruc, 1988; Syahrum, Kamaludin, dan Arjatmo,1994 ; Sugiyanto, 1996). (1) Pengenalan (kontak Awal) Sperma dan Sel Telur. Untuk mengetahui kontak awal antara sperma dan sel telur, terlebih dahulu diketahui cara fertiliasinya. Pada hewan yang hidup di perairan, sebagian besar melakukan fertilisasi eksternal, artinya sel telur dan sperma sama-sama dikeluarkan di dalam air dan keduanya akan bertemu dengan mekanisme tertentu. Ada 2 mekanisme fertilisasi eksternal yaitu a. Daya tarik spesies spesifik sperma Daya tarik ini adalah semacam kemotaksis dimana sel telur dapat mengeluarkan suatu zat yang mempunyai daya tarik (atraktan). Saat ini telah ditemukan 2 macam atraktan pada spesies spesifik seperti speract dari Strongylocerntrotus purpuratus dan resactdari Arbicia punctulata. Zat-zat ini merupakan polipeptida pendek dan terdapat di dalam selaput lendir telur. b. Reaksi akrosoma Reaksi akrosoma ini merupakan interaksi yang kedua antara sperma dan sel telur menyangkut aktivasi sperma oleh selaput lendir telur. Reaksi akrosoma ada 2 bagian 1. Gelembung akrosoma pecah mengeluarkan enzim pencerna. 2. Pembentukan prosessus akrosoma Kedua bagian tersebut menyebabkan kepala sperma dapat menembus selaput lendir dan prosessus akrosoma akan mencapai membran telur. Protein dari prosessus akrosoma yang berperan dalam proses ini adalah bindin. Beberapa uji biokimia dari bindin menunjukkan bahwa bindin dari masing masing species dari satu genus
sangat berbeda, karena pada membran vitelin sel telur terdapat reseptor yang species specifik bagi bindin. Jadi pada tahap ini terjadi pengenalan atau kontak awal yang bersifat species spesifik. Pada hewan mamalia, sebagian besar melakukan fertilisasi internal artinya fertilisasi terjadi di dalam saluran genital betina bagian anterior, sehingga proses fertilisasi disesuaikan dengan lingkungannya. Mekanisme fertilisasi pada mamalia ada 3 perioda: a. Perioda Kapasitasi Perioda ini terjadi setelah terjadi ejakulasi sperma untuk waktu cukup lama di dalam saluran reproduksi betina, dan waktu ini untuk masing-masing species berbeda. Sperma yang baru diejakulasi belum mampu melakukan reaksi akrosoma. b. Perioda enzimatik perioda ini terjadi setelah terjadi reaksi akrosoma pada sperma, maka sperma dapat mengkonsentrasikan enzim proteolitiknya yaitu akrosin (enzim hialuranidase)secara langsung pada tempat dimana sperma terikat (Bruc, 1988) c. Perioda pelekatan sperma dan sel telur. Menurut (Syahrum, Kamaludin, dan Arjatmo, 1994) pada perioda ini terjadi antara sel telur dan sperma sama-sama menghasilkan zat yang saling memberi pengaruh satu sama lain yaitu ginogamon(dikeluarkan oleh sel telur) dan androgamon(dikeluarkan oleh sperma). Ada 2 macam ginogamon yaitu ginogamon 1 fungsinya mempercepat jalannya sperma dan ginogamon 2 yang menyebabkan permukaan ovum mudah melekatkan kepala sperma. Proses pelekatan antara sperma dan sel telur dapat terjadi dengan bantuan enzim akrosin dan ginogamon 2.
(2) Pengaturan Pemasukkan Sperma ke Sel Telur, pengaturan tersebut mulai dari masuknya kepala sperma ke dalam telur biasanya Monospermi artinya hanya satu sperma yang masuk ke dalam telur. Pada proses ini satu sperma bersatu dengan nukleus haploid telur, agar kromosom yang normal bagi suatu spesies dapat dicapai kembali. Namun bila sperma yang masuk lebih dari satu, disebut Polispermi, dan keadaan ini abnormal, umumnya mati atau tidak terbentuk zigot (Bruc, 1988). Kebanyakan hewan akan melakukan pencegahan agar polispermi tidak terjadi dengan melalui 2 cara yaitu dengan pencegahan cepat (temporer) dan lambat (permanen) (Sudarwati,S; L.A. Sutasurya 1994). Pencegahan secara cepat terjadi dalam waktu 2-3 detik dan berlangsung selama 60 detik. Depolarisasi membran terjadi karena ada influks Na+ ke dalam sel, sehingga mengakibatkan terjadi perubahan potensial membran. Pencegahan secara lambat ditandai dengan terjadi reaksi korteks dan didahului dengan proses demobilisasi Ca 2+ ke daerah tempat masuknya sperma. Reaksi korteks mulai dengan pecahnya granula korteks dan dengan cepat granula melepaskan isinya berupa cairan ke dalam rongga yang mengelilingi telur sehingga terbentuk rongga privitelin.Cairan tersebut oleh granula korteks akan melarutkan ikatan antara membran vitelin dengan membran korteks. Pada waktu yang sama air akan masuk dan melepaskan membran vitelin, dan membran vitelin yang lepas disebut membran fertilisasi (Sudarwati,S; L.A. Sutasurya 1994;Sugiyanto, 1996). Di alam terdapat berbagai cara untuk mencegah polispermi. Pada Mamalia, kemungkinan terjadi polispermi sangat kecil dengan sedikitnya sperma yang dapat mencapai tempat fertilisasi (Bruc, 1988). Pada telur-telur yang mengandung banyak yolk seperti pada burung, reptilia, dan salamander, beberapa sperma sebenarnya masuk ke dalam telur,
namun dengan mekanismeyang belum diketahui semua sperma yang masuk mengalami disintegrasi di dalam sitoplasma, sehingga hanya satu sperma yang dapat melebur dengan pronukleus. Pada saat sperma masuk atau penetrasi ke dalam sel telur yang tidak kuat akan tertinggal dan mati pada corona radiata, dan hanya satu yang dapat menembus zona pelusida (Syahrum, Kamaludin, dan Arjatmo,1994; Sugiyanto, 1996). Disini terlihat bahwa hanya satu sperma haploid yang dapat berfusi dengan nukleus haploid telur. C. PENUTUP Uraian mengenaiawal terjadi fertilisasi meliputi mekanisme kontak awal antara sperma dan sel telur, serta mekanisme yang cukup kompleks dengan mengkaji mengenai proses pengaturan pemasukkan sperma ke dalam sel telur. Pada proses ini menunjukkan bahwa hanya satu sperma saja yang dapat menembus membran sel telur, meskipun pada keadaan tertentu ada kemungkinan beberapa sperma mampu menembus beberapa sel telur. C. PERTANYAAN/TUGAS 1. Mengapa anda mempelajari fertilisasi pada struktur perkembangan hewan II ? 2. Jelaskan maksud dengan pengenalan atau kontak awal yang bersifat species spesifik secara fisiologis pada fertilisasi eksternal. 3. Buatlah skema mekanisme fertilisasi eksternal, agar lebih mudah difahami! 4. Jelaskan 3 perioda mekanisme fertilisasi internal pada mamalia, mana paling lama? 5. Mungkinkah terjadi polispermi pada mamalia, jelaskan alasannya? TUGAS 1. Buatlah uraian singkat kecenderungan hanya satu sperma saja yang dapat menembus sel telur ?. 2. Setiap mahasiswa diwajibkan menyerahkan artikel ilmiah (dari jurnal/majalah
ilmiah/internet edisi 3 tahun terakhir) yang berkaitan dengan fertilisasi: monospermi/polispermi dan diserahkan setelah melalui diskusi kelompok paling lambat seminggu setelah ujian tengah semester. E. DAFTAR PUSTAKA 1. Bruc, M.C. 1988. Patten’s Foundations of Embryology. 5th ed. “gametogenesis and fertilization, cleavage and formation of the blastula” McGraw-Hill Book Company. New York San Fransisco Mexico Milan Montreal Panama Singapore Sydney Tokyo Toronto. Pp. 73—185 2. Syahrum; M. H Kamaludin; T Arjatmo. 1994. Reproduksi dan Embryologi . Dari Satu Sel Menjadi Organisme: Fertilisasi. Fakultas Kedokteran Universitas Indonesia, Jakarta. Hal. 63 – 87. 3. Sugiyanto, J. 1996. Perkembangan Hewan. Fakultas Biologi, Universitas Gajah Mada, Yogyakarta. Proyek Pendidikan Tenaga Akademik. Direktorat Jenderal Pendidikan tinggi, Departemen Pendidikan dan kebudayaan, Jakarta. Hal. 95—210. 4. Sudarwati,S; L.A. Sutasurya 1994. Dasar-dasar Struktur Perkembangan Hewan. Diktat kuliah Jurusan Biologi FMIPA- ITB. Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Teknologi Bandung. Hal.120 – 222
MODUL/TOPIK 6 FERTILISASIAKHIR A. PENDAHULUAN Modul 6 ini menjelaskan mengenai proses fertilisasi akhir yang membahas mengenai peleburan bahan genetik yang terjadi setelah masuknya kepala sperma ke dalam telur, yang dilanjutkan dengan proses aktivasi metabolik sel telur yang baru saja dibuahi. Pada proses ini satu sperma bersatu dengan nukleus haploid telur, agar kromosom yang normal bagi suatu spesies dapat dicapai kembali. Selain itu juga membahas mengenai adanya penyimpangan dari fertilisasi antara lain partenogenesis. Setelah mempelajari modul ini mahasiswa diharapkan mampu memahami tentang mekanisme dari akhir dari fertilisasi baik secara morfologis, chemis, dan fisiologis terutama saat terjadi peleburan bahan genetik dan aktivasi metabolik dari sel telur. Kemudian mahasiswa juga dapat memahami penyimpangan proses fertilisasi melalui diskusi kelompok . B. URAIAN Pada akhir dari proses fertilisasi menurut Sudarwati dan Sutasurya (1994) meliputi peleburan bahan genetik yang terjadi setelah kepala sperma masuk ke dalam sel telur. Kemudian diteruskan dengan proses aktivasi metabolik telur yang membran intinya akan melakukan disintegrasi. Bahan inti berinteraksi dengan sitoplasma telur, dan kromatin
mulai meregang. Menjelang berakhirnya peregangan kromatin membran intiyang baru mulai terbentuk dan membran ini disebut pronukleusjantan. Bersamaan dengan proses terbentuknya pronukleus jantan, granula mitokondria dan aparatus golgi darisperma akan menyebar di dalam sitoplasma sel telur dan inti telur, kemudian akan menjadi haploid yang disebut pronukleus betina (Gillbert,1994). Dari bagian sperma lainnya hanya sentriol yang dipertahankan dan akan menjadi aster. Aster adalah semacam bintang yang muncul pada saat terjadi pembelahan sel baik secara mitosis maupun meiosis, dan aster ini berada di ujung kutub animalis dan kutub vegetatuvus (Gillbert,1994). Aster tersebut berperan penting dalam mendekatkan pronukleus jantan dan pronukleus betina. Dengan bantuan aster dari sperma, pronukleus betina bergerak ke bagian tengah telur dan mendekati pronukleus jantan. Setelah kedua pronukleus bertemu, kedua membran pronukleus tersebut akan melakukan proses peleburan, yang dalam hal ini akan menyatukan 2 kromosom paternal dan kromosom maternal di dalam satu membran. Setelah peleburan pronukleus terjadi, DNA kromosom berreplikasi sebagai persiapan pembelahan pertama dari zigot. Pada proses replikasi tersebut diawali dengan tersusunnya kromosom pada keping metafase yang dilanjutkan proses anafase dan telofase, maka proses fertilisasi telah berakhir dan zigot siap memasuki masa perkembangannya atau proses embriogenesis (Bruc, 1988;. Sudarwati dan Sutasurya 1994) . Selain terjadi peleburan sperma dan sel telur dalam fertilisasi, ada keadaan lain dari sperma dan sel telur yang tidak fertil yang disebut infertilitas. Faktor-faktor yang mempengaruhi proses infertilitas atau sterilitas antara lain disebabkan oleh kelainan genetik atau kelainan
hormonal. Selain itu juga disebabkan oleh penyakit yang mengganggu fungsi organ seks, misalnya alkoholisme, gangguan endokrin, peradangan, penutupan saluran kelamin yang disebabkan oleh penyakit gonorehoe, dan tuberculosis. Selain factor penyakit dan gangguan fungís organ, infertilitas juga dapat terjadi oleh faktor kejiwaan seperti ejaculatio precox pada pria, atau disebut dengan sex maniak (Syahrum, dkk.1994). PARTENOGENESIS Menurut Bruc (1988) partenogenesis adalah proses dari telur yang tidak difertilisasi dapat diaktivasi dan berkembang menjadi individu normal atau dengan kata lain proses fertilisasi tanpa kehadiran pejantannya. Kondisi ini terjadi pada beberapa kelompok hewan Invertebrata dan beberapa spesies Vertebrata. Partenogenesis biasanya terjadi pada kondisi lingkungan yang tidak normal antara lain kemarau panjang atau musim dingin atau salju (Bruc, 1988; Gillbert,1994). Partenogenesis dapat dilakukan secara buatan di laboratorium melalui beberapa cara. Cara yang paling sederhana adalah memasukkan jarum yang telah dicelup di dalam darah katak lalu di masukkan atau disuntikkan ke dalam telur katak. Apabila dalam penyuntikan tidak tepat, dengan kata lain pada kutub vegetativus, maka telur katak tersebut akan bereaksi dengan mengeluarkan darah tersebut dari dalam telur, sehingga bila diamati akan terjadi telur di dalam genangan darah (Gillbert,1994). Meskipun penyuntikan darah berhasil ditembuskan dari kutub animalis, namun sebagian besar telur yang dirangsang untuk partenogenesis sebagian gagal untuk hidup normal. Hal ini mungkin disebabkan oleh terbebasnya gen-gen resesif yang mengganggu di dalam tubuh embrio dari hasil partenogenesis. Sebagian lagi yang mampu hidup normal biasanya embrio yang diploid, hal ini mungkin karena badan polar ke duanya
dipertahankan didalam telur. Pada umumnya individu hasil dari partenogenesis adalah betina (Bruc, 1988). C. PERTANYAAN/TUGAS 1. Jelaskan arti peleburan bahan genetik dan proses aktivasi metabolik? 2 Jelaskan mengapa sampai terjadi infertilitas, faktor apa saja yang mempengaruhinya? 3. Dalam kondisi seperti apa hewan tertentu dapat melakukan partenogenesis? 4. Tuliskan proses partenogenesis pada 3 hewan Invertebrata dan 2 hewan Vertebrata lengkap dengan rincian prosesnya. TUGAS 1. Kaitkan kegiatan fertilisasi akhir dengan pembelahan mitosis dan meiosis! 2. Diskusikan mengapa terjadi infertilitas atau sterilitas pada hewan dan manusia. bahan diskusi bisa dari bahan ajar, literatur, dan internet. Tugas dikumpulkan sebelum ujian tengah semester D. PENUTUP Pada proses akhir dari fertilisasi adalah peleburan bahan genetik. Selain itu juga terjadi penyimpangan proses fertilisasi seperti partenogenesis. Dalam proses ini sperma dan sel telur yang bergabung masing-masing mempunyai kekurangan, tetapi saling komplementer. E. DAFTAR PUSTAKA 1. Bruc, M.C. 1988. Patten’s Foundations of Embryology. 5th ed. “gametogenesis and fertilization, cleavage and formation of the blastula” McGraw-Hill Book
Company. New York San Fransisco Mexico Milan Montreal Panama Singapore Sydney Tokyo Toronto. Pp. 73—185 2. Gillbert, S.F. 1994. Developmental Biology. 4th ed. Sinauer Associates Inc., Sunderland, Massachusetts. USA. Pp.121 –198. 3. Syahrum; M. H Kamaludin; T Arjatmo. 1994. Reproduksi dan Embryologi . Dari Satu Sel Menjadi Organisme: Fertilisasi. Fakultas Kedokteran Universitas Indonesia, Jakarta. Hal. 63 – 87. 4. Sudarwati,S; L.A. Sutasurya 1994. Dasar-dasar Struktur Perkembangan Hewan. Diktat kuliah Jurusan Biologi FMIPA- ITB. Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Teknologi Bandung. Hal.120 – 222 5. Sugiyanto, J. 1996. Perkembangan Hewan. Fakultas Biologi, Universitas Gajah Mada, Yogyakarta. Proyek Pendidikan Tenaga Akademik. Direktorat Jenderal Pendidikan tinggi, Departemen Pendidikan dan kebudayaan, Jakarta. Hal. 95—210.
MODUL/TOPIK 7 EMBRIOGENESIS: PEMBELAHAN ATAU SEGMENTASI A. PENDAHULUAN Embriogenesis adalah perkembangan setelah terjadi zigot dari hasil fertilisasi. Ada 2 cara perkembangan dari zigot yaitu secara pembelahan (segmentasi) dan blastulasi dan akan dilanjutkan dengan perkembangan embryo secara gastrulasi. Pada proses pembelahan sangat dipengaruhi oleh tipe telur dan juga jumlah yolk, sehingga terjadi tipe pembelahan secara holoblastik dan meroblastik. Selain tipe pembelahan tersebut, bentuk pembelahan akan bersifat radial dan bilateral simetris. Pada blastulasi tidak terjadi pertumbuhan, sebaliknya pada gastrulasi terjadi perubahan bentuk embrio dan terjadi pertumbuhan embrio. Setelah mempelajari modul ini mahasiswa diharapkan mampu memahami tentang mekanisme, macam, dan pembentukan alur-alur pembelahan atau segmentasi dari beberapa hewan baik Invertebrata maupun Vertebrata melalui diskusi kelompoknya. Selain itu mahasiswa juga akan mampu menjelaskan proses-proses pembelahan yang bersifat radial dan bilateral simetris pada beberapa hewan. B. URAIAN 1. Mekanisme Pembelahan
Proses pembelahan atau segmentasi, menurut Weichert (1970) dan Bruc (1988) adalah proses pada setiap organisme multiseluler yang memperbanyak diri melalui pembelahan sel-selnya, karena di dalam tubuh organisme itu sendiri mempunyai keterbatasan fungsi dalam melaksanakan kegiatan hidupnya, dimana sel-sel itu muncul melalui proses mitosis maupun meiosis. Pembelahan mitosis dimaksudkan untuk membangun tubuh, sedangkan pembelahan meiosis untuk membentuk membentuk generasi berikutnya. Organisme itu dibangun dari satu sel yaitu sel telur yang sudah dibuahi yang disebut zigot, melalui serangkaian pembelahan mitosis yang berjalan cepat dan perkembangan selnya. Ciri umum dari pembelahan zigot (Bruc, 1988; Gillbert, 1994) adalah 1.Zigot yang bersifat uniseluler akan diubah dengan adanya pembelahan mitosis yang berangsur-angsur akan membentuk yang multiseluler 2.Tidak terjadi pertumbuhan 3.Bentuk embrio tidak berubah, kecuali dengan terbentuknya suatu rongga di dalam embrio yang disebut blastosoel. 4.Tidak terjadi pembelahan kuantitatif dalam komposisi kimia telur, meskipun transformasi cadangan makanan menjadi sitoplasma yang aktif dan substansi sitoplasma menjadi substansi inti yang berlangsung. 5.Bagian-bagian sitoplasma telur tidak terjadi perubahan posisi secara mencolok dan pada umumnya tetap pada posisi yang sama seperti halnya dalam telur pada permulaan pembelahan. 6.Rasio inti sitoplasma yang pada permulaan pembelahan sangat rendah tetapi pada akhir pembelahan rasionya menjadi seperti yang terdapat pada sel somatis biasa. Seperti pada mitosis umumnya, pembelahan dari sel telur yang sudah dibuahi. Mula- mula terjadi pembelahan inti yang diikuti dengan pembelahan sitoplasma. Sel anak
yang terbentuk disebut blastomer. Dan sel-sel ini selanjutnya akan membelah lagi menjadi 4 blastomer, 8 blastomer, 16 blastomer, 32 blastomer dan seterusnya. Mula-mula pembelahan ini terjadi secara simultan pada semua blastomer, tetapi kemudian sinkronisasiini menghilang dan blastomer membelah pada waktu yang berlainan, bebas satu terhadap yang lainnya. Pembelahan blastomer ini merupakan pembelahan khusus dan struktur kromosomnya seperti pada sel somatis. Ada perbedaan yang penting antara pembelahan mitosis pada perkembangan akhir organisme dewasa dengan mitosis selama pembelahan pada perkembangan awal. Pada organisme dewasa pembelahan sel tersebut sangat erat hubungannya dengan tumbuh, artinya setiap kali membelah, sel-sel anak akan tumbuh sampai ukurannya menjadi dua kali lipat dari sel yang baru membelah, dan kondisi ini selalu terjadi saat pembelahan berikutnya. Namun sel-sel tersebut akan selalu mempertahankan ukurannya saat membangun jaringan (Sudarwati dan Sutasurya, 1994). Berbeda dengan perioda pembelahan awal zigot, pembelahan yang terjadi secara berurutan dari masing-masing blastomer tidak dipisahkan oleh perioda tumbuh. Satu blastomer tidak bertambah ukurannya menjelang pembelahan berikutnya (Gillbert, 1994). Akibatnya adalah pada setiap pembelahan blastomer yang terbentuk ukurannya hanya setengah dari sel asal. Pembelahan dimulai dengan sebuah sel yang besar dan berakhir dengan jumlah sel yang masing-masing tidak lebih besar ukurannya dari sel jaringan dewasa. Kenyataannya pada akhir pembelahan sel-sel biasanya justru lebih kecil dari sel-sel yang telah berdiferensiasi seluler yang sering disertai dengan bertambah besarnya ukuran masing-masing sel.
2. Faktor-faktor yang mempengaruhi pembelahan Ritme pembelahan yang dihitung oleh waktu antara dua mitosis tidak sama pada beberapa hewan. Kecepatan pembelahan ini nampaknya bergantung pada suhu, walaupun pada dasarnya tetap bergantung pada faktor genetik. Pada mamalia, sel somatis yang membelah cepat untuk membentuk suatu populasi tertentu membutuhkan waktu 15-20 jam pada suhu 37oC. Pada telur bulu babi, dapat mencapai blastula ± 1000 sel (10 generasi) dalam waktu beberapa jam pada suhu yang jauh lebih rendah (Syahrum, dkk.1994; Gillbert, 1994). Selain suhu ada faktor lain yang mempengaruhi pembelahan, sehingga menyebabkan tipe pembelahan yang berbeda-beda. Faktor-faktor tersebut seperti adanya tipe telur yang berdasarkan jumlah yolk a.Oligolesital adalah telur yang mempunyai sedikit yolk, misalnya: pada amphioxus dan mamalia. b.Mesolesital adalah telur dengan yolk sedang, misalnya pada Amphibia. c.Polilesital adalah telur yang mempunyai banyak yolk, misalnya pada Aves, reptilia, dan mamalia yang bertelur. Faktor lain adalah dengan penyebaran yolk, maka telur dapat dibedakan menjadi a.Isolesital yaitu telur dengan yolk yang tersebar merata, misal pada mamalia. b.Sentrolesital yaitu telur yang yolknya terdapat pada bagian tengah, seperti pada insekta. c.Telolesital yaitu telur yang yolknya terdapat pada salah satu kutub dan biasanya kutub vegetativus, misal pada Aves.
Telur atau zigot membelah menjadi blastomer yang masing-masing dipisahkan oleh bidang yang dapat disetarakan dengan bidang geometrik. Telur dapat dianggap sebagai bentuk bola bumi yang mempunyai 2 kutub yaitu kutub animalis dan kutub vegetativus(Weichert, 1970; Bruc, 1988; Gillbert, 1994). 3. Alur Pembelahan Bidang meridional yang melalui sumbu panjang telur nantinya akan menjadi bidang median embrio disebut bidang sagital. Bidangekuatorialmerupakan bidang yang dibuat sesuai dengan alur keliling telur yang membagi telur menjadi bagian yang masing-masing kutub yang berbeda. Letak alur itu tepat di ekuator telur atau tepat di tengah jarak kutub animalis dan kutub vegetativus. Bidang ini juga tegak lurus sumbu animal-vegetal. Bidang ekuator ini jarang tepat terjadi pada posisinya, karena seringkali bidang ini menggeser ke arah kutub animalis atau ke kutub vegetativus. Bidang vertikal merupakan bidang yang mengikuti alur tegak lurus pada sumbu bidang animal – vegetal atau bidang yang sejajar dengan bidang ekuatorial mendekati kutrub animal atau kutub vegetativus (Weichert, 1970). Berdasarkan alurnya pembelahan pada beberapa hewan ada 2 jenis yaitu 3.1 Pembelahan Holoblastik Menurut (Sudarwati dan Sutasurya, 1994) dan Sugiyanto (1996) dengan adanya perbedaan distribusi dan jumlah yolk, maka terdapat perbedaan dalam pembentukan alur pembelahan. Alur ini muncul sebagai suatu cincin di sekitar sel. Cincin ini akan membagi sel menjadi 2 dengan suatu gerakan dari semua arah ke bagian dalam sel. Pada telur oligolesital, seluruh sel akan terbelah secara sempurna menjadi 2 blastomer, karena alur pembelahan ini memotong seluruh bagian telur, dan pembelahan ini disebut holoblastik. Pembelahan ini dapat terjadi secara ekual atau secara tidak ekual.
3.1.1 Pembelahan holoplastik secara ekual Pembelahan ini biasanya terjadi pada Amphioxus atau pada katak. Pada Amphioxus, alur bidang pembelahan pertama dan melalui bidang meridional, dan pembelahan kedua melalui bidang meridional lagi dan posisinya tegak lurus dengan bidang pertama. Pembelahan ketiga adalah bidang ekuatorial yang sedikit ke arah kutub animalis, sehingga blastomer di bagian polus animalis lebih kecil dari pada blastomer dibagian kutub vegetativus. Blastomer kecil disebut mikromer, sedang yang besar disebut makromer. Letak mikromer tergeser 45o atau di bagian atas antara 2 mikromer yang berdekatan. Pembelahan keempatadalah 2 bidang meridional yang saling tegak lurus (Bruc, 1988; Syahrum, dkk. 1994). Pembelahan kelima adalah pada 2 bidang latitudinal, satu bidang di bagian polus animalis, dan 4 yang lain di bagian kutub vegetativus. Mikromer yang paling dekat dengan polus animalis berukuran paling kecil, sebaiknya di dekat kutub vegetativus makromer yang paling besar dan paling banyak mengandung yolk. Bidang keenam adalah meridional yang terjadi secara simultan yang menghasilkan 64 sel. Ketika terjadi bidang pembelahan ke delapan, sudah terbentuk blastosoel yang berisi cairan. Tiap-tiap blastomer tidak ada perbedaan pigmen (Sugiyanto, 1996). Cara pembelahan yang serupa dengan pada amphioxus adalah pada telur kelinci. Bidang pembelahan pertama meridional dan membentuk 2 blastomer yang bilateral simetris. Bidang pembelahan kedua juga meridional yang memotong tegak lurus bidang pertama. Kedua blastomer pertama mengandung belahan gray crescent yang sama. Pada tingkat 4 sel, kedua blastomer tidak mengandung belahan gray crescent.
Bidang pembelahan ketiga adalah latitudinal yang mengahsilkan 4 mikromer dan 4 makromer. Bidang pembelahan ke 4 adalah 2 bidang meridional, alur bidang pembelahan pada permulaannya hanya terjadi pada kutub animalis. Sampai pembelahan ke empat ini masih serupa dengan pada Amphioxus dan mulai pembelahan ke lima, mikromer akan membelah lebih cepat dari makromer. Mikromer lebih banyak mengandung pigmen hitam (Gillbert, 1994; Sudarwati dan Sutasurya, 1994). 3.1.2 Pembelahan holoplastik tidak ekual Pembelahan ini terjadi pada mamalia dan akan menghasilkan blastomer yang tidak sama besar. Pada kebanyakan telur mamalia, tingkat dua sel terdiri dari blastomer kecilsebagai sel formatif dan blastomer besar sebagai calon jaringan tambahan atau tropoblast. Pada telur mamalia, stadium dua sel , bagian kutub animalis terdapat blastomer berukuran kecil, sedangkan pada kutub vegetativus terdapat blastomer yang berukuran besar. Polosit mamalia terletak diantara ke dua blastomer tersebut, sebaliknya untuk polosit pada chordata rendah, cenderung merupakan indikasi bagian kutub animalis. Mikromer pada chordata rendah lebih cepat membelah, sebaliknya pada mamalia yang cepat membelah adalah makromernya. Pada stadium 2 sel, makromer membelah lebih dahulu sampai menjadi stadium 3 sel, kemudian diikuti oleh mikromer menjadi stadium 4 sel. Demikian seterusnya pembelahan selalu berurutan menjadi stadium 5 sel, 6 sel, 7 sel, 8 sel, dan seterusnya. Blastula mamalia disebut blastosis dan membentuk blastocoel disebabkan oleh pergantian tropoblast (Sudarwati dan Sutasurya, 1994; Sugiyanto, 1996). 3. 2 Pembelahan Meroblastik Beberapa telur Telolesital seperti pada amphibia, pembelahannya secara holoblastik, tetapi yolk berpengaruh pada kecepatan pertumbuhan alur. Pada katak, yolk terkonsentrasi pada
kutub vagitatus pembelahan terjadi secara bertahap pada bagian ini. Akibatnya pada kutub animalis akan terbentuk sel yang lebih banyak, maka pembelahan terbatas pada suatu diskus, yaitu sitoplasma yang tidak mengandung yolk disebut blastodiskus. Pada tipe telur ini alur pembelahan tidak sempurna dan pembelahannya disebut meroblastik (Bruc, 1988; Syahrum, dkk. 1994) Pembelahan meroblastik menghasilkan sel-sel yang akan tumbuh embrio yaitu yang terletak di tengah atau disebut sel formatif. Sel-sel yang akan menjadi selaput embio berasal dari sel-sel marginal. Pembelahan terjadi di sekitar inti dan ooplasma yang berbentuk cakram yang disebut blastodis. Bidang pembelahan pertama adalah miridional yang memotong blastodis, tetapi tidak pernah mencapai kutub vegetativus. Bidang pembelahan kedua juga meridional dan tegak lurus pada bidang pertama. Bidang pembelahan ketiga meridional di kanan kiri bidang ke dua. Pembelahan keempatadalah vertikal yang sejajar dengan bidang pertama dan memotong bidang kedua. Setelah bidang keempat pembelahan menjadi tidak teratur lagi, kemudian diorientasikan ke dalam 3 pola pembelahan, yaitu: (a) Terjadi pada alur pembelahan vertikal yang melingkar (radier). (b) Memotong alur-alur bidang radier, sampai terjadi sel baru dan menambah jumlah sel sentral. (c) Bidang latitudinal atau horizontal memotong bidang vertikal, sehingga terbentuk sel permukaan dan sel bagian bawah. Batas sel-sel yang jelas hanya terjadi pada permukaan saja, sedangkan di bagian bawah tidak terdapat batas-batas yang jelas dan sitoplasma saling berhubungan. Setelah terbentuk bidang horizontal dan terbentuk 60 – 100 sel, maka terdeapat sel-sel sentral (sentroblast) dan sel periblast. Diantara sentroblast dan periblast terdapat blastosoel.
Pada telur sentrolesital, pembelahan inti mula-mula terjadi pada bagian tengah telur dan tidak disertai denga pembelahan sitoplasma. Akibatnya akan dihasilkan sejumlah nukleus yang terdapat pada bagian tengah sel. Setelah beberapa kali, inti mulai bergerak keluar ke arah permukaan. Jika inti sudah mencapai permukaan telur, sitoplasma yang mengelilinginya bercampur dengan sitoplasma dari lapisan permukaan (Syahrum, dkk. 1994; Sudarwati dan Sutasurya,1994; Sugiyanto, 1996). Pada tahap berikutnya, sitoplasma membelah dan alur pembelahan bergerak dari permukaan ke bagian dalam sebanyak inti yang dibentuk, dan bagian ini yang kemudian disebut sel, meskipun pada awalnya sel ini masih berhubungan dengan yolk.Akhirnya sel-sel ini terpisah sama sekali dari yolk dankemudian yolk ini merupakan massa padat dan berguna sebagai sumber makanan bagi embrio. Tipe pembelahan ini disebut superfisial Pembelahan telur menjadi blastomer, biasanya sangat teratur dan pola pembelahannya ditentukan oleh susunan aparatus mitosis. Aparatus mitosis ini bergantung pada susunan sitoplasma yang diprogram selama oogenesis, sehingga pembelahan pertama biasanya vertikal melalui sumbu utama telur. Bidang pembelahan kedua juga vertikal juga melalui sumbu utama tetapi tegak lurus terhadap bidang pembelahan pertama. Bidang pembelahan ketiga tegak lurus terhadap bidang pembelahan pertama dan kedua, juga terhadap sumbu utama. Oleh sebab itu pembelahan ini termasuk horizontal atau paralel terhadap ekuator telur. Hasil dari pembelahan ini adalah terbentuk 8 sel dengan 4 sel terletak di bagian atas, dan yang 4 sel lagi terletak dibagian bawah. Pada pembelahan radial tiap-tiap blastomer dari deretan bagian atas letaknya sesuai dengan blastomer dari deretan bagian bawah,
sehingga dihasilkan embrio yang radial simetri (Syahrum, dkk. 1994; Sudarwati dan Sutasurya,1994; Sugiyanto, 1996). Pada pembelahan secara spiral, blastomer dari deretan bagian atas terletak pada bagian atas dari batas antara dua blastomer dari deretan bagian bawah. Sususnan demikian disebabkan oleh kumparan pembelahan arahnya miring. Keempat kumparan pada pembelahan ketiga tersusun bentuk spiral, yang arah spiral jika dilihat dari atas sesuai dengan arah jarum jam dan sebaliknya. Jika arah spiral sesuai dengan jarum jam disebut dekstral, jika arah spiral bertentangan dengan jarum jam disebut sinistral (Sudarwati dan Sutasurya, 1994; Sugiyanto, 1996)) Semua pembelahan dari tipe spiral ini siklus pembelahannya berganti-ganti, atau kalau satu pembelahan searah dengan jarum jam, maka pembelahan selanjutnya akan berlawanan arah. Tipe pembelahan secara keseluruhan, baik dekstral maupun sinistral bergantung pada arah spiral yang terjadi pada pembelahan ketiga. Pada Lymnea (keong sawah) suatu lokus gen bertanggung jawab terhadap pembelahan ini. Baik dekstral atau sinistral dapat ditemukan pada jenis keong ini, tetapi yang dekstral lebih dominan. Oleh sebab itu kalau induk homozigot sinistral dikawinkan dengan jantan homozigot dekstral, semua keturunannya langsung sinistral. Hal ini disebabkan oleh induknya yang mempunyai gen sinistral, sehingga oosit dibentuk dibawah panduan gen sinistral. Akibatnya sitoplasma diprogram untuk membelah sinistral. Genom sperma disini tidak berpengaruh karena sudah ditentukan pada pembelahan awal (Gillbert, 1994).
Pembelahan yang hampir menyerupai pembelahan radial adalah tipe bilateral. Pada stadium 4 sel, 2 buah blastomer mempunyai ukuran lebih besar dari lainnya. Pada pembelahan ini, bagian kiri dan kanan embrio serupa dan dipisahkan oleh suatu bidang yang disebut bilateral simetri. Kegiatan pembelahan pada satu bagian merupakan cermin dari bagian lainnya. Salah satu contoh pembelahan bilateral pada nematoda, Pada pembelahan pertama terbentuk 2 sel dan salah satu selnya mempunyai ukuran lebih besar. Sel yang lebih besar diberi tanda AB, dan sel yang lebih kecil ditandai dengan P1. Kemudian sel-sel ini membelah lagi, sel AB menjadi sel A dan sel B, sedangkan sel P1 akan membelah menjadi P2 dan EM St, sehingga stadium ini menyerupai huruf T (Sugiyanto, 1996). Susunan seperti ini hanya untuk sementara, karena kemudian sel-sel P2 bergerak ke arah sel B. Pada pembelahan ketiga sel A dan sel B membelah ke kanan dan ke kiri, sedangkan sel lainnya membelah ke arah depan dan belakang, selanjutnya sel-sel tersebut akan membelah lagi menjadi: 1. Sel A menjadi sel α dan sel a 2. Sel B menjadi sel β dan sel b 3. sel P2 dan M St menjadi sel P3, C, E, dan Mst. Pembelahan pada nematoda ini juga merupakan contoh pembelahan determinate, dimana suatu blastomer membentuk bagian tertentu dari embrio yaitu: 1. Blastomer A, B, C membentuk kulit dan saluran pencernaan makanan 2. Blastomer E membentuk endokrin 3. Blastomer M St membentuk mesoderm dan Stomodeum 4. Blastomer P3 membentuk sel-sel reproduksi Yolk yang terdapat di dalam telur sejak mulai pembelahan berpengaruh pada prosesnya. Komponen sel seperti kromosom, bagian-bagian sitoplasma, mitokondria dan lapisan permukaan sel, semuanya turut aktif dalam pembelahan sel, sedangkan yolk bersifat pasif.
Jika jumlah yolk ini terlalu banyak, maka dapat menghambat atau menghalangi proses pembelahan. Akibatnya blastomer yang menerima yolk lebih banyak, pembelahannya lebih lambat dan ukurannya lebih besar dibandingkan dengan blastomer yang menerima yolk lebih (Gillbert, 1994; Syahrum, dkk. 1994; Sudarwati dan Sutasurya, 1994). Pada telur katak, pengaruh yolk ini dapat dilihat pada pembelahan pertama. Selama anafase terbentuk alur pembelahan pada permukaan telur dan akan memisah menjadi 2 blastomer. Alur ini terbentuk tidak serentak di sekeliling telur tetapi hanya pada kutub animalis yang hanya berisi sedikit yolk. Secara bertahap akan terbentuk alur ke arah kutub vegetativus sampai akhirnya telur terpisah menjadi 2 blastomer. Proses yang sama terulang kembali pada pembelahan kedua. Pada pembelahan ketiga, bidang pembelahan arahnya horizontal, alur terbentuk simultan di sekeliling telur. C. PERTANYAAN/TUGAS 1. Benarkah pembelahan dari setiap embrio ditentukan oleh gen induk? Berikan bukti 2. Buatlah intisari dari ciri umum dari pembelahan zigot menurut Gillbert (1994) 3.Apakah faktor-faktor yang mempengaruhi pembelahan pada setiap hewan sama? Berilah contoh dan penjelasannya. 4. Mengapa terjadi perbedaan alur pembelahan pada masing-masing hewan? 5. Bandingkan antara pembelahan holoblastis dan meroblastis. 6. Jelaskan mekanisme pembelahan radial, spiral, dan bilateral pada hewan tertentu. dan dimana letak kesamaannya 7. Jelaskan hubungan antara pembelahan determinate dengan pembentukan fungsi organ TUGAS 1. Hubungkan bentuk pembelahan radial, spiral dan bilateral dengan kategori taksonomi. 2. Gambarkan hal prinsip dalam proses pembelahan atau cleavage dari beberapa hewan
baik Invertenrata maupin Vertebrata. Tugas harus didiskusikan antar kelompok dan laporan juga dalam kelompok yang harus diserahkan sebelum ujian tengah semester. D. PENUTUP Akhir dari pembahasan ini terlihat kompleksitas dari pembelahan atau segmentasi baik dari mekanisme, alur, tipe, dan hasilnya. Setiap hewan ada yang sama mekanisme, alur maupun tipe pembelahannya, akan tetapi banyak yang berbeda karena pengaruh dari faktor eksternal maupun internalnya. Bila mampu mengkaji kompleksitas segmentasi dengan cermat, maka akan menemukan pemahaman yang retensi. E. PUSTAKA 1. Bruc, M.C. 1988. Patten’s Foundations of Embryology. 5th ed. “gametogenesis and fertilization, cleavage and formation of the blastula” McGraw-Hill Book Company. New York San Fransisco Mexico Milan Montreal Panama Singapore Sydney Tokyo Toronto. Pp. 73—185 2. Gillbert, S.F. 1994. Developmental Biology. 4th ed. Sinauer Associates Inc., Sunderland, Massachusetts. USA. Pp.121 –198. 3. Syahrum; M. H Kamaludin; T Arjatmo. 1994. Reproduksi dan Embryologi . Dari Satu Sel Menjadi Organisme: Fertilisasi. Fakultas Kedokteran Universitas Indonesia, Jakarta. Hal. 63 – 87. 4. Sudarwati,S; L.A. Sutasurya 1994. Dasar-dasar Struktur Perkembangan Hewan. Diktat kuliah Jurusan Biologi FMIPA- ITB. Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Teknologi Bandung. Hal.120 – 222 5. Sugiyanto, J. 1996. Perkembangan Hewan. Fakultas Biologi, Universitas Gajah Mada, Yogyakarta. Proyek Pendidikan Tenaga Akademik. Direktorat Jenderal
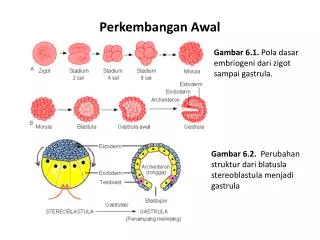
Pendidikan tinggi, Departemen Pendidikan dan kebudayaan, Jakarta. Hal. 95—210. 6. Weichert, C.K. 1970. Anatomy Of The Chordates: Early Development and Histogenesis. 4th ed. MC Graw-Hill Company. New York, St. Louis, San Fransisco, Toronto, London. Pp.52 – 63 MODUL/TOPIK 8 EMBRIOGENESIS: BLASTULASI A. PENDAHULUAN Proses blastulasi terjadi setelah proses segmentasi berakhir, kemudian akan terbentuk blastoderm.. Kemudian bagian tepinya terbentuk blastomer yang melekat erat pada yolk membentuk periblast, yang diduga berperan dalam memecah yolk untuk pertumbuhan embrio.Setelah mempelajari modul ini mahasiswa diharapkan mampu memahami tentang mekanisme dan macam-macam proses blastulasi. Mahasiswa juga diharapkan tahu faktor- faktor baik eksternal maupun internal yang mempengaruhi blastulasi. B. URAIAN Proses blastulasi ini terjadi jika proses segmentasi berakhir mulai dari kumpulan blastomer yang merupakan proses akhir dari segmentasi blastodiskus, kemudian akan terbentuk blastoderm atau termasuk dalam stadium 32 sel.Pada stadium 32 sel, bidang pembelahan
terbentuk pada bagian bawah dan paralel terhadap permukaan, sehingga terbentuk lapisan sel permukaan yang terletak pada bagian atas dari lapisan sel yang berhubungan dengan yolk. Sitoplasma yang terdapat pada kutub animalis, bagian tegaknya membelah menjadi sejumlah blastomer bebas. Kemudian bagian tepinya terbentuk blastomer yang melekat erat pada yolk dan merupakan lapisan yang saling berhubungan, membentuk kelompok sel yang disebut periblast. Periblast ini tidak berperan langsung dalam pembentukan tubuh embrio, tetapi diduga berperan dalam memecah yolk untuk pertumbuhan embrio (Bruc, 1988; Gillbert, 1994; Syahrum, dkk, 1994; Sudarwati dan Sutasurya 1994 ;Sugiyanto, 1996). Blastomer pada pembelahan awal berbentuk bulat seperti telur, karena tekanan permukaan blastomer yang saling bersentuhan menjadi rata, tetapi permukaan bebas nya tetap bundar, bentuk embrio pada stadium ini disebut morula. Penyusunan kembali blastomer dalam stadium morula dapat berbeda-beda tergantung dari kelompok hewannya. Berdasarkan atas bentuk dan susunan blastomernya, blastula dibedakan menjadi 3 macam yaitu: (1) Soelo blastula (blastula bundar) Bentuk seperti bola, berasal dari telur yang bertipe oligolesital dan mesolesital dengan pembelahan secara holoblastik teratur, misalnya terdapat pada Amphioxus dan Amphibia. (2) Disko blastula (blastula pipih) Bentuk seperti cakram, berasal dari telur yang bertipe oligolesital yang mengalami pembelahan secara holoblastik tidak teratur dan telur polilesital yang membelah secara meroblastik. Blastula terdapat di atas yolk atau jaringan penyalur makanan. Misalnya terdapat pada ikan, reptil, burung dan monotremata, yang menyebut blastula dengan germinal disk, yang terdiri dari jaringan embriodan periblast.Pada eutheria blastula
disebut dengan blastokist, yang mempunyai 2 kelompok sel yaitu embrioblast atau massa sel dalam (” inner cell mass ”) dan tropoblast (Bruc, 1988; Gillbert, 1994). (3) Stereoblastula (blastula bulat masif) Blastula berbentuk bulat seperti bola seperti pada soeloblastula tetapi lebih masif, yang terdapat pada Gymnophiona dan Ganoidea (Sugiyanto, 1996). Menurut Sudarwati dan Sutasurya (1994), pada prinsipnya blastula dari Chordata terdiri dari blastoderm dan blatocoel. Semua blastula seperti kuntum pada tubuh hewan yang akan membentuk bagian-bagian tubuh hewan, seperti: a. Neuroektodermal Daerah ini akan tumbuh dan berkembang menjadi sistem saraf b. Notokordal Daerah ini nantinya akan berkembang menjadi korda dorsalis c. Mesodermal Daerah ini biasanya sepasang, sehingga apabila terjadi pertumbuhan, akan menjadi organ-organ yang berpasangan d. Endodermal Daerah ini akan menjadi sistem pencernaan dan pernafasan e. Germinal Daerah yang mengandung plasma germinal yang akan menjadi sel kelamin induk f. Prekordal (lamina prekordalis) daerah ini akan menjadi organisator di daerah kepala Lokasi calon-calon itu diatur pola dari masing-masing golongan hewan. Calon itu masih pula diorganisasi dan penyusunan lokasi baru dalam proses gastrulasi. Reorganisasi atau penggeseran dan pergerakan menuju ke posisinya yang terakhir, sehingga perkembangan
menjadi organ dan calon satu terhadap yang lain akan saling berintegrasi. Bentuk blastula berhubungan erat dengan pola proses gastrulasi. Hal tersebut disebabkan oleh blastosoel yang memberi kesempatan kepada calon-calon organ menuju ke posisi dalam tubuh embrio. Tingkat blastula dalam perkembangan berperan untuk mempersiapkan susunan calon-calon yang disangka akan membentuk organ tubuh (Sugiyanto, 1996; Sudarwati dan Sutasurya 1994) . Blastomer bertambah besar, akan mempengaruhi bentuk, sifat maupun kelompok sel tertentu mempunyai struktur yang berbeda dengan keadaan semula. Potensi merupakan kemampuan sel baru yang spesifik dalam proses deferensiasi. Kemampuan sel untuk membentuk suatu calon organ berangsur-angsur menurun setelah proses pembelahan. Kemampuan pada stadium 2 sel lebih besar daripada stadium 4 sel, stadium 4 sel lebih besar dari stadium 8 sel dan seterusnya. Hal ini lebih jelas seperti pada Echinodermata, pada stadium 2 sel dapat terbentuk embrio yang lengkap, demikian pula pada stadium 4 sel. Pada stadium 8 sel keadaannya lain, blastomer di kutub vegetativus berkembang menjadi gastrula. Sedangkan totipotensilebih menyatakan tingkat perkembangan blastomer yang masih mampu berperan sebagai calon dari berbagai macam sel atau struktur yang akan terbentuk (Bruc, 1988; Sugiyanto, 1996). Dengan adanya proses perkembangan, maka akan terjadi pembatasan potensi. Suatu keadaan yang menyatakan bahwa dalam suatu tingk atau sebelum terjadi pembatasan potensi disebut pleuripoten (Sugiyanto, 1996). Determinasi suatu sel atau kelompok sel akan terarah dan hasil-hasilnya akan mempunyai penentu (determinant). Determinant dapat berupa bentuk, letak atau kandungan bahan yang ada dalam sel. Lebih spesifik lagi determinant berupa gen dalam kromosom. Potensi sel dari hasil proses determinasi adalah
terbatas, artinya sel tersebut tidak mampu lagi manjadi calon dari bermacam-macam jaringan. Pada tingkatan yang totipoten, maka tiap blastomer dapat berkembang menjadi embrio atau sampai terbentuk individu baru dalam tiap-tiap blastomer. Kembar untuk menyatakan terbentuknya 2 individu baru atau lebih. Menurut proses kejadiannya, maka kembar dibedakan menjadi kembar fraternal (dizigotik) dan kembar identik(monozigotik) (Syahrum, dkk, 1994). Pada individu baru yang terbentuk pada kembar fraternal berasal dari ovum yang berbeda. Supervolusi pada manusia dapat terjadi kembar apabila masing- masing ovum dibuahi. Pada kucing, anjing, atau babi, ovum yang masak secara berurutan dapat membentuk beberapa embio dalam satu kebuntingan. Tiap-tiap individu baru yang terbentuk dari satu ovum disebut kembar identik atau kembar sejati. Pemisahan yang sempurna dari blastomer akan terjadi pemisahan embio yang sempurna pula. Bila pemisahan tidak sempurna, maka akan terjadi kembar siam (dampit). Kembar monozigotik berkembang dalam satu bungkus embrio, sedangkan kembar fraternal berkembang dalam bungkus embrio yang terpisah (Bruc, 1988; Gillbert, 1994; Syahrum, dkk, 1994). C. PERTANYAAN 1. Jelaskan hal-hal yang sangat prinsip dan mekanisme pada proses blastulasi 2. Jelaskan mekanisme terjadi kembar identik atau kembar sejati dan kembar siam. 3. Jelaskan faktor-faktor yang mendukung kelancaran proses blastulasi 4. Hubungkan proses blastulasi dengan pembentukan sistema organ hewan 5. Dimana kesulitan anda dalam memahami prinsip-prinsip blastulasi.
D. PENUTUP Pada proses blastulasi mekanismenya berbeda oleh susunan blastomernya. Semua blastula seperti kuntum pada tubuh hewan yang akan membentuk bagian-bagian dari suatu sistema. Pada proses determinasi suatu sel atau kelompok sel mempunyai penentu (determinant). Determinant yang spesifik lagi determinant berupa gen dalam kromosom. Sebenarnya bentuk blastula berhubungan erat dengan pola proses gastrulasi. E. PUSTAKA 1. Bruc, M.C. 1988. Patten’s Foundations of Embryology. 5th ed. “gametogenesis and fertilization, cleavage and formation of the blastula” McGraw-Hill Book Company. New York San Fransisco Mexico Milan Montreal Panama Singapore Sydney Tokyo Toronto. Pp. 73—185 2. Gillbert, S.F. 1994. Developmental Biology. 4th ed. Sinauer Associates Inc., Sunderland, Massachusetts. USA. Pp.121 –198. 3. Syahrum; M. H Kamaludin; T Arjatmo. 1994. Reproduksi dan Embryologi . Dari Satu Sel Menjadi Organisme: Fertilisasi. Fakultas Kedokteran Universitas Indonesia, Jakarta. Hal. 63 – 87. 4. Sudarwati,S; L.A. Sutasurya 1994. Dasar-dasar Struktur Perkembangan Hewan. Diktat kuliah Jurusan Biologi FMIPA- ITB. Fakultas Matematika dan Ilmu Institut Teknologi Bandung. Hal.120 – 222 5. Sugiyanto, J. 1996. Perkembangan Hewan. Fakultas Biologi, Universitas Gajah Mada, Yogyakarta. Proyek Pendidikan Tenaga Akademik. Direktorat Jenderal Pendidikan tinggi, Departemen Pendidikan dan kebudayaan, Jakarta. Hal. 95—210. Institut Teknologi Bandung. Hal. 120 – 222