Download

1 / 15
190 likes | 511 Vues
Explore the complex interplay of insulin and glucagon in regulating glycogen metabolism and glucose homeostasis. Learn about phosphorylation cascades, organ specialization, and metabolic consequences.

E N D
Review pathways and organ specialization • Phosphorylation cascades in glycogen phosphorylase and glycogen synthase regulation • Glucagon and epinephrine signalling through cAMP-dependent protein kinase (PKA) • Glycolysis vs. gluconeogenesis • Regulation of GLUT4 (glucose transport) • The insulin receptor is a tyrosine kinase (RTK) • Next time: Insulin signalling through phosphoinositols and MAPK’s, connection to obesity Regulation by Insulin and Glucagon Jason Kahn: Insulin and Glucagon
Overview of Metabolic Regulation • We need to understand how metabolic regulation is supposed to work before we can understand how it fails in diabetes. • See Voet and Voet 3/e, Chapter 27 and chapters 19-26. Magisterial but dense. • Flux through pathways is determined at a few key steps. • Levels of critical metabolites are highly regulated, largely allosterically: much more rapid than gene expression. • Blood glucose is the critical mis-regulated metabolite in diabetes. Many downstream consequences such as glycalation and ketoacidosis. Jason Kahn: Insulin and Glucagon
Organ Specialization • Insulin signals fed state: largely controls removal of glucose from plasma • Many agents, including glucagon and epinephrine, stimulate mobilization. • Hence they act differently on different tissues. For example, in the liver cAMP stimulates glycogen breakdown and activates gluconeogenesis (in order to export glucose), but in heart muscle cAMP stimulates glycogen breakdown and glycolysis, to increase energy production. Jason Kahn: Insulin and Glucagon
Insulin and Glucagon • Insulin: Signals fed state. • Secreted by pancreatic cells when blood glucose is high. • Stimulates glucose uptake by muscle and other tissues. • Stimulates glycogen synthesis, fatty acid synthesis, protein synthesis. • Represses gluconeogenesis and glycogen breakdown. • Upregulates genes for fatty acid biosynthesis. • Acts through a receptor tyrosine kinase that turns on many pathways in the cell. • Glucagon: Signals hungry state. • Secreted by pancreatic cells when glucose is low. • Represses glycogen synthesis and fatty acid synthesis. • In the liver, stimulates gluconeogenesis and glycogen breakdown, leading to glucose export. • Upregulates gene for PEPCK. • Acts through a G-protein coupled receptor that activates adenylate cyclase. • Diabetes is caused by either lack of insulin (Type I) or lack of proper response (Type II), leading to increased blood glucose because uptake is not turned on. Jason Kahn: Insulin and Glucagon
Glycogen Metabolism • Glycogen is a polymeric form of glucose, with (14) linkages and (16) branches. Similar to starch. Cellulose has (14) linkages, giving sheet structures that are much harder to break down. Primarily found in muscle and liver. • Glycogen is synthesized by glycogen synthase, from glucose-6-phosphate via phosphoglucomutase and UDP-glucose pyrophosphorylase. Glycogen synthesis is energetically uphill. • Glycogen is broken down by glycogen phosphorylase and debranching enzyme; this is exergonic in the cell. It is degraded starting from an end, so the branched structure means that glucose can be released rapidly. Debranching is slower. • Since glycogen is broken down from the ends, the branched structure provides rapid glucose release • Glycogen synthesis and are catalyzed by different enzymes, so they can be independently regulated and do not occur simultaneously in the same tissue. Jason Kahn: Insulin and Glucagon
Regulation of Glycogen Metabolism I • V+V 3/e chapter 18 • We will work backwards. • The choice between glycogen synthesis (glycogen synthase) and breakdown (glycogen phosphorylase) is mediated mainly by phosphorylation and dephosphorylation of the two enzymes. • Terminology:The “a” form is more active than the “b” form.The “o” form is the original, the “m” form is modified.The “R” quaternary state is more active than the “T” state (Glycogen synthase kinase 3 = GSK-3 Jason Kahn: Insulin and Glucagon
Regulation of Glycogen Metabolism II • Coordinated regulation: phosphorylase is activated by phosphorylation, synthase is inhibited. • Balance of activities is therefore determined by the balance between activities of phosphorylase kinase and phosphoprotein phosphatase I Active • Phosphorylase a is more active because it is preferentially in the “R” form, but it can still be converted to the “T” state by high glucose. Phosphorylase b usually remains in the “T” state at physiological [ATP] Inactive Who will regulate the regulators? Jason Kahn: Insulin and Glucagon
Regulation of Glycogen Metabolism III • Phosphorylase regulation is a classic bicyclic enzyme cascade. This allows for signal amplification because every phosphorylation event by the first kinase creates an active enzyme that acts on the downstream target. • Protein kinase A, or cyclic AMP dependent protein kinase, is activated by dissociation of its regulatory subunit. • So…how is [cAMP] regulated? Jason Kahn: Insulin and Glucagon
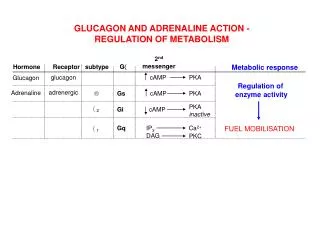
Control of Adenylate Cyclase • Adenylate cyclase (AC) is activated or inactivated by G-proteins that are in turn activated by transmembraneG-protein coupled receptors or GPCRs (about which more later). • Glucagon is a small (29-residue) peptide hormone that binds the glucagon receptor, primarily found in the liver and skeletal muscle. • Binding of glucagon to its receptor leads to activation ofadenylate cyclase: the receptor catalyzesexchange of GTP forGDP, hence activatingthe G protein. • cAMP is the classic“second messenger,” asmall molecule thattransduces a signal fromoutside the cell to inside. • cAMP phosphodiesteraseshuts off the signal. A number of mechanisms for the effects of caffeine have been suggested. Caffeine stimulates the release of intracellular calcium and inhibits the activity of cyclic nucleotide phosphodiesterases at concentrations between 0.5 and 1 mM. These concentrations are well above those seen in the blood in response to normal intakes of caffeine suggesting that the effects of caffeine are mediated by other mechanisms. It is now generally accepted that physiological concentrations of caffeine (about 100 μM) act by antagonising the effects of adenosine. Caffeine acts at A1 adenosine receptors which are negatively linked to adenyl cyclase and A2a adenosine receptors which are positively linked to adenyl cyclase. Hence, competition between caffeine and adenosine at cell surface A1 and A2a adenosine receptors leading to changes in the intracellular concentration of cyclic AMP is the likely mechanism underlying the physiological effects of caffeine. Jason Kahn: Insulin and Glucagon http://www.cosic.org/background-on-caffeine
Additional Layers I • Phosphorylase kinase is also activated by Ca++, via calmodulin (its subunit PhK ). Since Ca++ release from intracellular stores is the signal for muscle contraction, this coordinates glycogen breakdown with demand for ATP. • Phosphoprotein Phosphatase 1 (PP1) was the first serine-threonine phosphatase discovered. It dephosphorylates phosphorylase kinase, glycogen phosphorylase, and glycogen synthase, thus stimulating glycogen synthesis. Its substrate enzymes are also glycogen-bound. • PP1c (catalytic) is bound to glycogen via its GM (muscle) or GL (liver) regulatory subunit, or else bound to PP1-inhibitor 1 (or other inhibitor proteins), preventing it from acting as a “loose cannon” when glycogen is depleted. • PP1-Inh 1 is activated by phosphorylation by PKA, so glucagon stimulation of PKA leads to increased inhibition of PP1c as well as increased stimulation of phosphorylase kinase • Is anyone bothered by the idea that PP1-Inh 1 appears to be both a substrate and an inhibitor of PP1c? Cohen (EJB, 1983) suggests that it binds the active site so as to block other substrates but allow its own dephosphorylation Jason Kahn: Insulin and Glucagon
Additional Layers II • Remember that PP1c is bound to GM, which is itself activated by phosphorylation by an insulin-stimulated protein kinase, leading to glycogen synthesis. • But dual phosphorylation of GM (by, who else?, PKA) releases PP1c and allows its inhibition, so that the bicyclic kinase cascade signal is maintained. • In the liver, GL also stimulates PP1c, but GL-PP1c is allosterically inhibited by m-phosphorylase a, the form that is active in glycogenolysis. The “T” state of phosphorylase a is induced by high glucose, and the T state (not the R state) is a substrate for PP1c. This give o-phosphorylase b, which dissociates. However, phosphorylase is in 10-fold excess over PP1c in the cell, so PP1c is not free from inhibition until it has dephosphorylated most of the phosphorylase. • Get it? This creates a time delay, so that glycogen synthase cannot be dephosphorylated (activated) until glycogen phosphorylase has already been substantially dephosphorylated, and this only occurs in the presence of glucose. Don’t stomp on the brakes until after you have released the accelerator! Glycogen synthase itself is also a better substrate for PP1c when it has bound substrate G6P. Reset by phosphatases PP2A and PP2B (calcineurin) Jason Kahn: Insulin and Glucagon
Effects of Insulin So Far • Stimulates PP1c via phosphorylation of GM by ISPK (via IRS1, Insulin receptor substrate 1). A similar kinase activates protein biosynthesis (S6 kinase). • Inhibits kinases that inactivate glycogen synthase. • Here’s more: Jason Kahn: Insulin and Glucagon
Glycolysis vs. Gluconeogenesis • In glycolysis, fructose-6-phosphate is phosphorylated by phosphofructokinase 1 (PFK-1) to give fructose-1,6-bisphosphate. • In gluconeogenesis, the reverse transformation is simply hydrolysis, catalyzed by fructose bis-phosphatase 1 (FBPase-1) • The two enzymes are allosterically regulated by fructose-2,6-bisphosphate, which is made and destroyed by separate PFK-2 and FBPase-2 activities respectively. • F-2,6-BP activates PFK-1 and inhibits FBPase-1, throws the switch toward …glycolysis. (PFK-1 is also allosterically inhibited by ATP and activated by AMP.) • PFK-2 and FBPase-2 are two activities resident on one protein! • In the liver, phosphorylation by…PKA inhibits PFK-2 and activates FBPase-2, favoring gluconeogenesis • In heart muscle, a different PFK-2/FBPase-2 isozyme responds oppositely, favoring glycolysis Jason Kahn: Insulin and Glucagon
Role of Insulin in Glucose Transport • Glucose transporters act as gated pores. Glucose is retained in the cell by phosphorylation to give G6P. • In the absence of insulin, muscle cells are impermeable to glucose. • Liver doesn’t listen to insulin: it responds directly to glucose concentration, and is permeable to glucose in order to either ship it out or sock it away. • Liver buffers high glucose by using a low-affinity version of hexokinase (glucokinase), which imports glucose at concentrations above 5 mM. • Insulin induces glucose uptake by promoting exocytosis of the GLUT4 transporter from vesicles to the plasma membrane. The transporter is subsequently recycled back. • This is mediated by the IRS-1 docking protein and its associated PI-3 kinase, which localize on the vesicle surface. • Additional mechanisms for GLUT4 exocytosis operate through caveolae, membrane domains related to rafts enriched in signalling proteins. Jason Kahn: Insulin and Glucagon
Allosteric Regulation of Glycolysis • The PFK and PK steps of glycolysis are inhibited by ATP and activated by AMP. • AMP is a better sensor than ADP for the energy charge ([ATP]+0.5*[ADP])/([ATP]+ [ADP]+[AMP]), because of the adenylate kinase reaction:2ADP <=> ATP + AMP • Small changes in [ATP]thus lead to large changes in [AMP] relative to its generallyvery small value. Jason Kahn: Insulin and Glucagon