Download
1 / 26
270 likes | 797 Vues
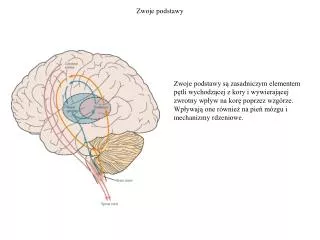
Zwoje podstawy. Zwoje podstawy są zasadniczym elementem pętli wychodzącej z kory i wywierającej zwrotny wpływ na korę poprzez wzgórze. Wpływają one również na pień mózgu i mechanizmy rdzeniowe. Zwoje podstawy.

E N D
Zwoje podstawy Zwoje podstawy są zasadniczym elementem pętli wychodzącej z kory i wywierającej zwrotny wpływ na korę poprzez wzgórze. Wpływają one również na pień mózgu i mechanizmy rdzeniowe.
Zwoje podstawy W skład zwojów podstawy wchodzi kilka połączonych ze sobą struktur: jądro ogoniaste (caudate) i skorupa (putamen) (tworzących razem prążkowie (striatum)), gałka blada (globus pallidus), istota czarna (substantia nigra) i niskowzgórze (subthalamic nucleus).
Zwoje podstawy Połączenia anatomiczne ukazujące równoległe bezpośrednie i pośrednie drogi z prążkowia do jąder wyjściowych zwojów podstawy. Dwa rodzaje receptorów dopaminergicznych znajdują się w dwóch grupach neuronów prążkowia początkując drogi bezpośrednie i pośrednie. Drogi hamujące zaznaczone na szaro, drogi pobudzające, na różowo. GPe = external segment of the globus pallidus; GPi = internal segment of the globus pallidus; SNc = substantia nigra pars compacta; STN = subthalamic nucleus. Droga bezpośrednia tworzy sprzężenie zwrotne dodatnie, a pośrednia sprzężenie zwrotne ujemne w obwodzie pomiędzy zwojami podstawy i wzgórzem. Aktywacja drogi bezpośredniej usuwa hamowanie wzgórza powodując wzrost jego aktywności. Aktywacja drogi pośredniej zwiększa hamowanie wzgórza. Aktywacja drogi bezpośredniej zwiększa aktywność ruchową, aktywacja drogi pośredniej hamuje ruchy.
Zwoje podstawy Obwody zwojów podstawy w warunkach normalnych, chorobie Parkinsona, hemibalizmie (hemiballism, niekontrolowane wymachy kończynami) i pląsawicy (chorea - niekontrolowane i nieregularne ruchy). Drogi hamujące zaznaczone na szaro i czarno, drogi pobudzające, na różowo i czerwono.Degeneracja substancji czarnej w chorobie Parkinsona powoduje zmiany w obu projekcjach z prążkowia zaznaczone zmianą koloru (ciemniejszy odcień oznacza wzmożoną aktywność). W chorobie tej wyjście hamujące do wzgórza jest zwiększone, a w hemibalizmie i pląsawicy jest zmniejszone. GPe = external segment of the globus pallidus; Gpi = internal segment of the globus pallidus; SNc = substantia nigra pars compacta; STN = subthalamic nucleus.
Choroba Parkinsona – terapie chirurgiczne Wycięcie niskowzgórza (lewo) lub wewnętrznej części gałki bladej są efektywnym sposobem redukcji objawów choroby Parkinsona.
Choroba Parkinsona – terapie lekowe Ilustracja choroby ParkinsonaA Manual of Diseases of the Nervous System,1886 Levodopa jest naturalnym aminokwasem przekształcającym się w mózgu w dopaminę poprzez dekarboksylację. L-dopa przechodzi przez barierę krew-mózg w przeciwieństwie do dopaminy. Karbidopa i entakapon hamują enzymy rozkładających L-dopę w obwodowym układzie nerwowym i zwiększają dostępność levodopy w mózgu. L – levadopa, C – carbidopa, E - entacapon Książka Olivera Sacksa i film „Przebudzenie” opowiadające o doświadczeniach z 1969 r. w leczeniu katatonii przy użyciu L-dopy.
Choroba Parkinsona – deep brain stimulation Operacja implantacji elektrody do głębokiej stymulacji mózgu. Stymuluje się globus pallidus interna (Gpi) i subthalamic nucleus (STN).
Kora ruchowa Kora ruchowa stanowi zasadniczy element kontroli ruchowej średniego poziomu. Lokalizacja kory u różnych ssaków, w tym u małp (1870), potwierdziła lokalizacje kory i sekwencję obszarów (nogi, ręce, twarz) przewidzianą przez Jacksona. Motoryczny homunkulus otrzymany przez Penfielda i Rasmussena (1950) metodą stymulacji kory mózgowej.
Trzy obszary kory ruchowej Kora ruchowa znajduje się w płacie czołowym (frontal lobe), tuż przed bruzdą centralną (Central Sulcus) nazywaną też bruzdą Rolanda (Rolandic fissure) i jest podzielona na trzy wzajemnie połączone obszary, pierwszorzędową korę ruchową (MI), dodatkową korę ruchową (SMA) i korę przedruchową (PM). SMA i PM stanowią wspólnie drugorzędową korę ruchową (MII). Funkcjonalnie oznacza to wielokrotną reprezentację mapy ruchowej w korze mózgowej.
Stymulacja pierwszorzędowej kory ruchowej A. Stymulacja magnetyczna kory motorycznej lub odcinka szyjnego rdzenia kręgowego (cervical spine) aktywuje drogę drogę korowo-rdzeniową i wywołuje odpowiedz EMG mięśni. B. Zapisy z kontralateralnych mięśni ramienia i dłoni wykazują wcześniejszą odpowiedź na stymulacje rdzenia niż na stymulacje kory. Załamek S jest artefaktem związanym z przyłożonym impulsem magnetycznym. Pierwszorzędowa kora ruchowa kontroluje proste własności ruchu.
Elektryczna stymulacja pierwszorzędowej kory ruchowej Dokładniejsze studia za pomocą mikrolektrod implantowanych (intracortical microstimulation or ICMS) pokazały, ze neurony w pierwszorzedowej korze ruchowej (MI) aktywują nie tylko pojedyncze mięśnie oraz że te same mięśnie są aktywowane przez wiele różnych obszarów kory. Na wykresach widać miejsca w MI, stymulacja których wywołuje aktywność EMG (wskazując na połączenie monosynaptyczne) w mięśniu ramienia (deltoid muscle) i nadgarstka (extensor carpi radialis; ECR). Mapy topograficzne (1/próg stymulacji) ukazują zarówno redundancje jak i przekrywanie się reprezentacji. Umożliwia to aktywacje różnych grup mięśni w różnych zadaniach.
Obszary przedruchowe - połączenia U naczelnych istnieją cztery główne obszary przedruchowe. A. Wejścia do kory pierwszorzedowej u małpy: PMd (dorsal premotor area); PMv (ventral premotor area),S1 (primary sensory cortex), SMA (supplementary motor area.), CMA (cingulate motor area). B. Wejścia do kory przedruchowej: pola 5 i 7 Brodmana, kora przedczołowa (prefrontal cortex, pole Brodmana 46). Obszary 5 i 7 Brodmana zajmują się integracją multisensoryczną w celu planowania ruchu. Obszar 46 zajmuje się pamięcią operacyjną (working memory) i przetrzymuje w pamięci informacje o lokalizacji przestrzennej obiektów, w czasie wykonania ruchu. Występują również gęste połączenia pomiędzy obszarami przedruchowymi, nie pokazane na rysunku.
Połączenia z móżdżkiem izwojami podstawy Kora przedruchowa i pierwszorzędowa kora ruchowa dostają również wejścia ze zwojów podstawy i móżdżku poprzez wzgórze. Zwoje podstawy i móżdżek nie wysyłają bezpośrednio połączeń do rdzenia kręgowego.
Plastyczność somatotopicznej organizacji kory ruchowej Organizacja kory ruchowej jest plastyczna i zmienia się pod wpływem uczenia i uszkodzeń. A. Widok normalny somatotopicznej organizacji kory ruchowej szczura z obszarami reprezentującymi mięśnie łap przednich, wąsów i okołooczne. B. Po przecięciu połączeń nerwowych z wąsami obszary mięśni łap przednich i okołooczne są zwiększone kosztem obszaru wąsów. Reorganizacja może się odbyć nawet w czasie paru godzin. Prawdopodobnie jest ona wynikiem utraty wejść sensorycznych z wąsów do kory ruchowej.
Plastyczność somatotopicznej organizaji kory ruchowej cd Ruchy zamierzone ulegają poprawie wraz z praktyką. B. Skany fMRI ukazujące aktywacje kory MI podczas wykonywania sekwencji ruchowej ćwiczonej przez 3 tygodnie 20 min. dziennie (prawo) oraz nowej sekwencji ruchowej (lewo). Reprezentacja wyćwiczonej sekwencji jest większa niż sekwencji kontrolnej. W eksperymencie tempo wykonania zadania było w obu sytuacjach takie samo.
Komórki V warstwy kory motorycznej dają początek drodze piramidowej Porównanie sześciu warstw kory w różnych obszarach mózgu. W V warstwie kory motorycznej widoczne ciała neuronów komórek Betza. Komórki Betza znajdują się głównie w obszarze ruchowym nogi. Z lewej: siedem warstw kory motorycznej wg. klasyfikacji Cajala. Warstwy są wyróżnione na podstawie względnej ilości dużych ciał neuronów (komórki piramidalne) i małych ciał neuronów (komórki piramidalne, gwiaździste i ziarniste). Z prawej: komórka Betza kory motorycznej obrazowana metodą Golgiego.
Równoległe drogi ruchowe Pośrednie i bezpośrednie połączenia kory ruchowej i pnia mózgu z boczną i przyśrodkową częścią rdzenia kręgowego. Bezpośrednia droga korowo-rdzeniowa (droga piramidowa, corticospinal tract) omija wybrzuszenie piramidowe w pniu mózgu i schodzi do pnia mózgu, gdzie tworzy połączenia polisynaptyczne (u wyzszych ssaków) lub monosynaptyczne (u naczelnych) z motoneuronami.85 % włókien przekracza linię środkową i tworzy boczną drogę korowo-rdzeniową, pozostałe aksony tworzą grzbietową drogę korowo-rdzeniową. Drogi pośrednie: droga czerwienno-rdzeniowa i droga korowo-siatkowo-rdzeniowa.
Równoległe drogi ruchowe Przecięcie drogi korowo-rdzeniowej powoduje jednostronną słabość w mięśniach u małpy. Ustępuje ona po upływie paru miesięcy i zwierzęta odzyskują normalne funkcje ruchowe dzięki przejęciu funkcji przez drogi pośrednie. Jednakże indywidualne ruchy palców są utracone na zawsze.
Kodowanie siły mięśni Częstość odpalania neuronów w korze ruchowej koduje siłę mięśni. A1,2 - dwa rodzaje zachowania neuronów w korze ruchowej podczas momentu siły przyłożonego do nadgarstka. B. W obu rodzajach neuronów częstość odpalania wzrasta (liniowo) z siłą mięśni. (torque – ang. moment obrotowy)
Kodowanie populacyjne kierunku ruchu Kierunek ruchu jest kodowany w korze motorycznej poprzez populacje neuronów. (A) Wytrenowana małpa porusza ręką w ośmiu różnych kierunkach. (B) Aktywność neuronu ruchowego w korze podczas ruchów w ośmiu kierunkach. C.Krzywa strojenia neuronu. D. Wiele neuronów korowych jest aktywnych w trakcie ruchu w danym kierunku. Kierunek każdej linii reprezentuje preferowany kierunek danej komórki, a długość linii jest proporcjonalna do częstotliwości jej wyładowań. Wypadkowy ‘wektor’ aktywności (czerwona strzałka) koduje wykonywany kierunek ruchu (czarna strzałka) (Georgopoulos, 1982)
W ruchach generowanych wewnętrznie, przed ruchem obserwowany jest ujemny potencjał w EEG w obszarze dodatkowej kory ruchowej (SMA) zanim nastąpi aktywacja pierwszorzędowej kory ruchowej. Jest to tzw. potencjał gotowości (ang. readiness potential RP lub niem. Bereitschaftspotential, BP), który sygnalizuje planowanie ruchu. W eksperymentach pokazano, że BP jest widoczny na ok. 400 ms przed zanim badany relacjonował świadom myślenie o wykonaniu ruchu. Potencjał przygotowujący Doświadczenie Libeta (1983): świadomość, że chce się ruszyć palcem początek BP
Obszary przedruchowe – funkcje Visual cue: małpa miała dotykać paneli na podstawie ich podświetlania w pewnej sekwencji. Prior training: dotykanie paneli wg. wcześniej wyuczonej sekwencji. Neurony w korze ruchowej MI odpalały przed i w trakcie ruchu, tak samo w obu zadaniach. Dodatkowa kora ruchowa (SMA) była aktywna tylko przed i w trakcie zadania wyuczonego. Boczna kora przedruchowa była aktywna tylko przed i w trakcie zadania wykonywanego na podstawie informacji wzrokowej. Wniosek: dodatkowa kora ruchowa programuje programy motoryczne wyuczonych sekwencji ruchowych. Boczna kora przedruchowa bierze udział w przygotowaniu i wykonaniu ruchu w odpowiedzi na bodziec zewnętrzny. Po długotrwałym treningu, aktywność w dodatkowej korze ruchowej zanika – sekwencje wykonywane automatycznie ‘przenoszą się’ do kory pierwszorzędowej.
Obszary przedruchowe Neuron w bocznej grzbietowej korze przedruchowej staje się aktywny gdy małpa przygotowuje się do ruchu w lewo. Instrukcja (podświetlenie jednego z czterech paneli) wskazuje panel do naciśnięcia po bodźcu wyzwalającym (zapalenie się diody). Odpalanie neuronu rozpoczyna się po pojawieniu się instrukcji i trwa do wykonania zadania po bodźcu wyzwalającym. Wniosek: kora przedruchowa bierze udział w przygotowaniu i wykonaniu ruchu w odpowiedzi na bodziec zewnętrzny (w tym doświadczeniu - wzrokowy). Zachodzi w niej uczenie asocjacyjne (Associative learning) pomiędzy bodzcem sensorycznym a dana reakcja ruchowa.
Transformacja sensomotoryczna Transformacja reprezentacji sensorycznej otoczenia na sygnały kontroli mięśni nazywa się transformacją sensomotoryczną. Oddzielne, równoległe drogi zapewniają transformację wzrokowo-motoryczną wymaganą do sięgania i chwytania. PMd – premotor dorsal, PMv – premotor ventral.
Komórki zwierciadlane Komórki w bocznej grzbietowej korze przedruchowej wykazują aktywność gdy małpa wykonuje zadanie lub gdy obserwuje wykonanie zadania. Komórki te są więc zaangażowane w abstrakcyjną reprezentację ruchu. A. Aktywność neuronu podczas obserwacji chwytania wykonywanego przez inna małpę. B. Aktywność neuronu podczas obserwacji chwytania wykonywanego przez eksperymentatora. C. Aktywność neuronu podczas chwytania wykonywanego przez małpę. Neurony te nazwano neuronami zwierciadlanymi (mirror neurons).
Obwody ruchowe - synteza • Ruch zamierzony rozpoczyna się ‘programem centralnym’, który aktywuje w odpowiedniej sekwencji obszary kory ruchowej. • Włókna korowo-rdzeniowe pobudzają motoneurony. • Motoneurony wywołują ruch mięśni. • Poprzez kolaterale, włókna korowo-rdzeniowe pobudzają komórki czuciowe (informacja o wysłanych komendach). • 5 i 6 Wyjścia czuciowe z mięśni, stawów i skóry dostarczają informacji zwrotnej o stanie skurczu mięśni i fazie wykonania ruchu.