Download
1 / 49
610 likes | 1.27k Vues
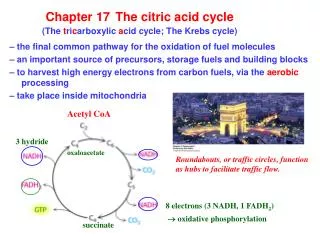
Chapter 17 The citric acid cycle (The t ri c arboxylic a cid cycle; The Krebs cycle). – the final common pathway for the oxidation of fuel molecules – an important source of precursors, storage fuels and building blocks

E N D
Chapter 17The citric acid cycle(The tricarboxylic acid cycle; The Krebs cycle) – the final common pathway for the oxidation of fuel molecules – an important source of precursors, storage fuels and building blocks – to harvest high energy electrons from carbon fuels, via the aerobic processing – take place inside mitochondria Acetyl CoA 3 hydride oxaloacetate Roundabouts, or traffic circles, function as hubs to facilitate traffic flow. 8 electrons (3 NADH, 1 FADH2) oxidative phosphorylation succinate
Coenzyme A: a carrier of acyl group thioester
2 m in length 0.5 m in diameter
The function of TCA cycle– the harvesting of high energy electrons from carbon fuels TCA + OP 90% energy production in aerobic cells No large amount of ATP generation No oxygen as a reactant (p. 490) Cellular respiration Substrate-level phosphorylation
§ 17.1 The link between glycolysis and the TCA cyclepyruvate + CoA + NAD+ acetyl CoA + CO2 + NADH • a specific pyruvate carrier embedded in membrane • decarboxylation and high-transfer-potential e- • Pyruvate dehydrogenase complex In mitochondria matrix
Pyruvate dehydrogenase complex (4~10 103 kd)–– increase the overall reaction rate and minimize side reactions Catalytic cofactors CoA, NAD+ Stoichiometric cofactors + Lys
Reactions of the pyruvate dehydrogenase complex order first
NAD+ NADH E1 C3 or C4 of Glc Derived by decarboxylation C C thiazole ring p.
Oxidation: E2 oxidized reduced Lipoyl-lysine arm of lipoamide
E2 Dihydrolipoyl transacetylase Activated acyl groups carrier Dihydrolipoyl dehydrogenase E3
Nelson Substrate channeling: all enzymes and coenzymes are clustered, allowing the intermediates to react quickly without diffusing away from the surface of the enzyme complex.
Citrate synthase:OAA + acetyl CoA + H2O citrate + CoA + H+ ¤synthase:two units are jointed without the direct participation of ATP ¤OAA binds first, induced a structure rearrangement, followed by acetyl CoA ¤ citryl CoA formation, thioester hydrolysis ¤CoA leaves the enzyme, followed by citrate, return to the original conformation Side effects p. 440
Aconitasecitrate cis-aconitate isocitrate An interchange of hydrogen atom and hydroxyl group hydration dehydration isomerase
Aconitase ¤ an iron-sulfur protein, or nonheme iron protein ¤ 4Fe-4S-3Cys, 1Fe binds to the carboxylate and hydroxyl groups of citrate ¤the availability of iron in the cell (02) An inhibitor of aconitase (2003) NTU Fluoroacetate (Garrette, p. 573)
Isocitrate dehydrogenase isocitrate + NAD+ -ketoglutarate +CO2 + NADH the determining rate of TCA cycle
-ketoglutarate dehydrogenaseis homologous to the pyruvate dehydrogenase complex Pyruvate + CoA + NAD+ acetyl CoA + CO2 + NADH
Succinyl CoA synthetase(succinate thiokinase) The only substrate-level phosphorylation in TCA cycle Thioester bond cleavage coupled to GDP phosphorylation E. coli enzyme uses either GDP or ADP as the phosphoryl-group acceptor Plants use ADP as the phosphoryl-group acceptor Nucleoside diphosphokinase: GTP + ADP GDP + ATP adenylate kinase
The committed point of TCA cycle– the allosteric enzymes Citrate synthase (in many bacteria), inhibit by ATP Citrate can be transported to the cytoplasm, inhibit glycolysis (phosphofructokinase) Key sites: Isocitrate dehydrogenase -ketoglutarate dehydrogenase
The source of biosynthetic precursors Acetate + CoA + ATP p. 460/493 Pyruvate carboxylase Acetyl CoA synthetase p. 495 (bacteria and plants, and humans) gluconeogenesis + AMP + PPi high / lowenergy charge Anaplerotic reaction
Glyoxylate cycle (in glyoxysomes of oil-rich seeds) Bypass two decarboxylation steps of TCA cycle isocitrate lyase and malate synthase (in bacteria and plants) 2 acetyl CoA + NAD+ + 2H2O succinate + 2 CoA + NADH + 2H+ TCA, gluconeogenesis (to power seeding growth) Metabolic versatility
ICL and MLS have as targets for therapeutic drugs to treat some bacterial and fungal infections
The Glyoxylate cycle —In plants, certain invertebrates, and some microorganisms acetate can serve both as an energy-rich fuel and as a source of phosphoenolpyruvate for carbohydrate synthesis — the enzymes are sequestered in the membrane-bound organelles, glyoxysomes, which are specialized peroxisomes — glyoxysomes develop in lipid-rich seeds during germination — glyoxysomes also contain all the enzymes needed for the degradation of fatty acids Nelson (05)
Beriberi ¤ a neurologic and cardiovascular disorder nervous system relies on Glc as its only fuel ¤a dietary deficiency of thiamine (vitamin B1) ¤ inactivated thiamine-related enzymes eg., pyruvate dehydrogenase, -ketoglutarate dehydrogenase, transketolase (a diagnostic indicator of red cells) ¤ limbs pain, musculature weakness, skin sensation disorder, heart enlargement, cardiac output inadequate ¤ similar symptoms for an organism is exposed tomercury orarsenite
Arsenite(AsO33-) poisoning– high affinity for neighboring sulfhydryls – lead to central nervous system pathologies (antidote) Arsenate (AsO43-) -mercaptoethanol Dithiothreitol (DTT) Dihydrollipoyl transacetylase Mad as a hatter: HgNO3 -amylase As2O3
Arsenate (AsO43-) glyceraldehyde 3-phosphate + Pi + NAD+ 1,3-bisphosphoglycerate + NADH + H+ 1-arseno-3-phosphoglycerate Ch.16 EX. 13
Citric acid/Citrate/citrate synthase: — three negatively charged carboxyl groups — a tart or fruity flavor — a plasticizer and foam inhibitor in some resins preparation, as a mordant to brighten colors, and as an antioxidant to preserve the flavors of foods — industrial production: Aspergillus niger, beet molasses as carbon source — a good chelator of metal ions — released into soil in some plants — alleviate the toxicity of Al3+ in acidic soil — highly expressed bacterial citrate synthase in plants Nelson (05) mg/g FW
Citric acid cycle: an amphibolic pathway, both catabolic and anabolic processes Nelson (05) ?
Regeneration of OAA ¤Succinate dehydrogenase:isoalloxazine ring of FAD ¤Fumarase:a stereospecific trans addition, L-isomer malate formation ¤Malate dehydrogenase:a significantly positive free energy ¤a metabolic motif: 2 oxidation, 1 hydration, methylene group (CH2) carbonyl group (CO) L- More energy is extracted in the form of FADH2 and NADH
Succinate dehydrogenase ¤ the free-energy change is insufficient to reduce NAD+ ¤ an iron-sulfur protein, contains three kinds: 2Fe-2S, 3Fe-4S, 4Fe-4S ¤ is embedded in the inner mitochondrial membrane,directly associated with the electron-transport chain
* 1953 Nobel * * * * * * * * * * * *
* * 2nd run * * * * * * * * * * * * * * * * * *
Ex. 11 and 12 p. 500 oxaloacetate Alexander (1948) an asymmetric enzyme which attacks a symmetrical compound can distinguish between its identical groups.
Acetyl CoA + 3 NAD+ + FAD + GDP + Pi + 2 H2O 2 CO2 + 3 NADH + FADH2 + GTP + 2 H+ + CoA Only under aerobic conditions Metabolon:the enzymes are physically associated with one another to facilitate substrate channeling between active sites.
Irreversible step p. 490 O2 does not participate directly in the TCA cycle. However, the cycle operates only under aerobic conditions. TCA energy generation
Regulation of pyruvate dehydrogenase complex ¤allosteric inhibitors: acetyl CoA (for E2), NADH (for E3) ¤reversible phosphorylation (for E1): energy charge, biosynthetic intermediates ¤ [Ca2+] cyto [Ca2+] mito activate phosphatase epinephrine (liver), [Ca2+] (p. 388) insulin (liver and adipose tissue), in fed state: glucose pyruvate acetyl CoA fatty acid synthesis The kinase is associated with the E2 The phosphatase is stimulated by Ca2+ A phosphatase deficiency: Glc lactic acid, unremitting lactic acidosis, central nervous system malfunction
At rest As exercise begins
§ 16.3 Regulation of the citric acid cycle Nelson (05) Allosteric effectors Covalent modification
揭開砒霜於白血病治療中的運作機制 作者:駐法科技組 97.05.09 摘要砒霜是治療某種罕見白血病十分有效的成份。位於Saint Louis醫院附設大學血液學研究院(Institut Universitaire d'Hématologie,混合機構分屬法國國家科學研究中心[CNRS]與巴黎第七大學)的研究員已經證實砒霜在罕見白血病治療中的機制。這些研究結果應該會讓我們進一步了解這類疾病的治療,進而找出更為有效的醫療策略。這項最新的研究成果由抗癌陣線所支持,同時已刊載於2008年4月13日自然細胞生物學(Nature cell biology)的線上期刊中。本文毒藥砒霜於醫界之運用已有三千多年的歷史。目前,砒霜在急性骨髓性白血病 (leucémie aiguë promyélocytaire)治療中經常可見。這種類型白血病的發展特徵在於骨髓性白血病(PML)蛋白質與RARA蛋白質的融合。PML-RARA蛋白質的融合就足以產稱白血病細胞。由Huges de Thé教授所率領的研究團隊首度發現砒霜會引起小泛素(SUMO)蛋白質的固定(fixation)。SUMO是一種肽,負責調節PML-RARA蛋白質間的互動。但這類複合體的降解(degradation)機制尚未解密,因為SUMO一般而言可以對抗蛋白質的降解。
法國研究員日前在降解機制中成功辨識RNF4酶。這種酶在經由砒霜修飾後的PML-RARA蛋白質形式的降解與辨識過程中,扮演著關鍵的角色。它會固定這個複合體(PML/RARA-SUMO)上的另一個泛素( ubiquitine)酶,而這種已知的酶將會引發其所接合的蛋白質降解。之後,泛素將針對PML/RARA-SUMO蛋白質進行修飾。這種由SUMO所引發並由泛素執行的降解管道的存在已經可以透過酵母(levure)的遺傳研究來預測,然而迄今尚未成功辨識出任何底物(substrat,或譯為受質)。目前的研究結果或許可以進一步了解這類疾病的治療,進而擬出更為有效的醫療策略。參考資料:Arsenic degrades PML or PML-RARA through a SUMO-triggered RNF4/ubiquitin-mediated pathway, Lallemand-Breitenbach, V., Jeanne, M., Benhenda, S., Nasr, R., Lei, M., Peres, L., Zhou, J., Zhu, J., Raught, B., and de The, H., Nature Cell Biology, en ligne le 13 avril 2008