Young Leaf
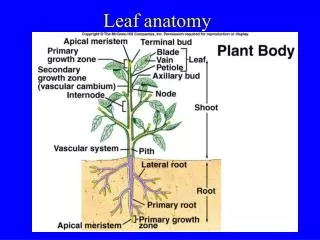
Young Leaf. Shoot Tip. Axillary Bud. Node. Internode. Phytomere Node Leaf Axillary Bud Internode. Increase in length of the stem occurs largely by internodal elongation. Node. Internode. Plant cells are surrounded by rigid cell walls. Cell migration does not occur in plants.

Young Leaf
E N D
Presentation Transcript
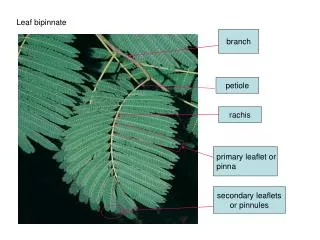
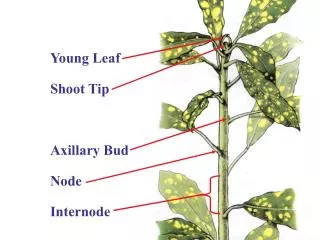
Young Leaf Shoot Tip Axillary Bud Node Internode
Phytomere Node Leaf Axillary Bud Internode
Increase in length of the stem occurs largely by internodal elongation. Node Internode
Plant cells are surrounded by rigid cell walls. Cell migration does not occur in plants.
Cell expansion plays a major role in growth Root cells expand their volume 50 times by expanding lengthwise but not widthwise
In roots, cell expansion plays a major role in growth Two competing plant hormones determine the direction of cell expansion: GA (gibberellic acid) promotes growth along the length Ethylene promotes growth along the width
Auxin and Cytokinin control shoot and root growth High levels of Cytokinin and low levels of Auxin promote shoot development (stems with leaves) High levels of Auxin and low levels of Cytokinin promote root development
The Miller-Skoog Experiment: Cloning • Place single cell on medium with high levels of Cytokinin and low levels of Auxin to promote shoot development (stems with leaves) • Place shoots on medium with high levels of Auxin and low levels of Cytokinin to promote root development at the base of the shoot • Transfer rooted shoots to soil and grow plants to maturity
The pollen “germinates” and grows down into the ovary where fertilization of the egg occurs.
A successful fertilization will produce a fertilized egg with 2X DNA.
Even at this one cell stage the embryo reveals polarity. The first cell division is asymmetric, producing a small apical cell and a larger basal cell.
The apical cell will later give rise to the entire “embryo proper”. The basal cell will give rise to a small umbilical cord-like structure called the suspensor.
The small apical cell divides several times to generate the globular embryo. All cells of this embryo appear morphologically similar.
Several divisions later morphological asymmetry is seen in the heart shaped embryo.
Gerd Jurgens searched for embryo pattern mutants. • Soak seeds in a mutagen • Grow plants to maturity • These plants would be carriers of mutations (m/+) • When these carriers self-fertilize, the resulting • embryos would be: +/+, m/+, m/m • Mutants similar to gap mutants in flies, lacking regions of the embryo, including the apical structures, the stem (hypocotyl) and root, were identified
L1 and L2 cells divide anticlinally: perpendicular to the surface These divisions contribute to surface growth without increasing the number of cell layers
L3 cells, or corpus layers, divide in both planes to add additional cell layers to the shoot.
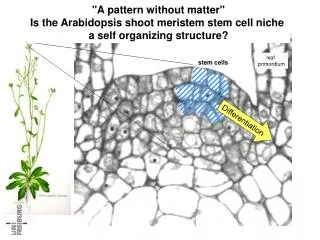
Shoot Apical Meristem The shoot apical meristem can be divided into distinct zones.
-stem cells Shoot Apical Meristem The central zone is maintained as a pool of undifferentiated stem cells.
-peripheral zone Shoot Apical Meristem The peripheral zone is the site of organ initiation.
-stem cells -peripheral zone Shoot Apical Meristem As cell divisions occur in the central zone, the resulting cells are pushed into the peripheral zone where they are incorporated into organ primordia.
-stem cells -peripheral zone Shoot Apical Meristem The central zone cells will give rise to all of the above-ground organs of the mature plant.
-stem cells -peripheral zone Shoot Apical Meristem How is the stem cell population maintained throughout the life of the plant?
-stem cells -peripheral zone Shoot Apical Meristem A feedback loop between organ initiation and the stem cell population regulates the size of the meristem.
Genes Controlling Meristem Development Normal heart- stage embryo WUS or STM mutant embryo WUSCHEL and SHOOTMERISTEMLESS mutants fail to develop a shoot apical meristem.
Genes Controlling Meristem Development Normal heart- stage embryo WUS or STM mutant embryo CLV1 or CLV3 mutant embryo CLAVATA1 and CLAVATA3 mutants develop a greatly enlarged shoot apical meristem.
Genes Controlling Meristem Development • STM and WUS are required to form and maintain the stem cell population • CLV1 and CLV3 are required to prevent the over-proliferation of the undifferentiated stem cell population
Genetic Interactions between STM and CLV • The greatly enlarged meristems that form in clv mutants are largely suppressed when the activity of STM is reduced (for example, in stm/+ plants).
Genetic Interactions between STM and CLV • The greatly enlarged meristems that form in clv mutants are largely suppressed when the activity of STM is reduced (for example, in stm/+ plants). • Similarly, the loss of shoot meristems in stm mutants is restored in plants that have reduced CLV activity (for example in clv/+ plants).
Genes Controlling Meristem Development • STM and WUS are homeobox genes and encode proteins that function as transcription factors • CLV1 encodes a receptor protein • CLV3 encodes a small protein that functions as a signaling molecule that binds to the CLV1 receptor
CLV / WUS Interactions CLV3 is expressed in the L1 and L2 cell layers of the central zone
CLV / WUS Interactions CLV1 and WUS are expressed in a small domain of L3 cells in the central zone
CLV / WUS Interactions CLV3 expression is lost in WUS mutants. Therefore, WUS activates CLV3 expression.
CLV / WUS Interactions The expression domain of WUS is greatly enlarged in CLV1 and CLV3 mutants. CLV3 binds to and activates the CLV1 receptor, which then represses WUS expression.
CLV / WUS Interactions WUS is part of an “organizing center” that promotes stem cell proliferation in overlying cells.
CLV / WUS Interactions A feedback loop between CLV and WUS maintains the stem cell population throughout the life of a plant.