Download

1 / 47
470 likes | 613 Vues
This article explores ion channels' crucial function in facilitating selective substance entry into cells, overcoming the limitations of the phospholipid bilayer's selectivity. It discusses the criteria for passage through the bilayer, the energetic challenges faced by ions, and how channels lower activation energy, similar to enzymes, to enable faster ion flow. The structure, function, and biochemical aspects of ion channels, including voltage-gated channels, are examined to understand their importance in cellular processes like action potentials.

E N D
The Cell Membrane is Selective • Criteria for passage through the phospholipidbilayer: • Hydrophobic • Net zero charge • Nonpolar • Size is also a consideration • Chemicals that will NOT pass through the phospholipidbilayer: • Hydrophillic • Charged, ionic • Polar • Size is also a consideration
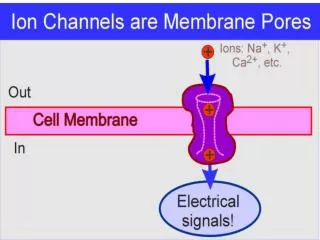
But, using the preceding criteria, many substances vital to cellular function (e.g., ions) and survival (e.g., glucose) will not gain entry into the cell! • So, nature has come up with channels, which selectively allow certain substances to gain entry into the cell, even though they do not meet the criteria on the preceding slide.
Channels are Vital Without channels it is energetically unfavorable to move ions across a membrane – 1. the phospholipidbilayer is ~6-8 nm thick. 2. the hydrophilic head of the phospholipid molecule projects toward the cytoplasm or the extracellular fluid. 3. the hydrophobic tails of the phospholipid molecules project toward each other.
For the Cation to Move Through the PhospholipidBilayer…, • It must lose its waters of hydration so that it is not so huge and charged; requires energy to break attractive forces between the ion and the waters. • Energy is also required to move a charged highly hydrophilic particle into the highly hydrophobic area of the lipid bilayer that contains the “tails” of the phospholipid molecules. • Based on thermodynamic calculations, so much energy would be required for this process that it would never occur.
Channels are thermodynamically similar to Enzymes in that the former lower the Activation Energy required to move ions across the Membrane Transition State The rate of the reaction is determined by the energy of activation, the energy input required to produce the transition state. The uncatalyzed reaction requires a higher activation energy than the catalyzed one does. So, the latter runs more quickly. There is no difference in free energy (ΔG) between uncatalyzed and catalyzed reactions. The ΔG is the thermodynamic driving force for the reaction and determines the direction of the reaction. A + B AB C + D Reactants transition products (substrates) state
Ion Flow Across the Membrane • A Chemical can move across the membrane through one of two ways: • Movement through the phospholipidbilayer. • Movement through a H2O-filled protein channel.
Channels are thermodynamically similar to Enzymes in that the former lower the Activation Energy required to move ions across the Membrane The rate of diffusion is determined by the “energy of activation”, the energy input required to produce the “transition state”. (Remove H2Os of hydration and/or move into hydro-phobic environ-ment.) Movement through a channel only requires shedding of waters of hydration (energy input would be infinite to move through the bilayer). So. Diffusion occurs more quickly. ΔG determines this!! Transition State Change in free energy is the thermodynamic driving force for diffusion and determines the direction of ion movement A + B AB C + D Reactants transition products (substrates) state
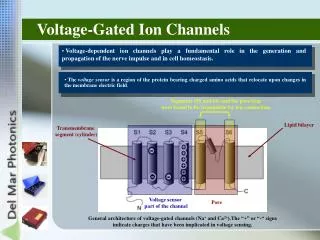
What do we know about the structure of gated ion channels? • Biochemical Information – • MWs range from 25-250 kDal. • They are integral membrane glycoproteins. • They usually consist of 2 or more subunits. • The genes that code for the proteins have been isolated, cloned and sequenced. These sequences have been grouped into 6-7 protein families. • The primary (amino acid) sequences of these channels is known.
Use of the Hydrophobicity Plots • Propose 3-D structures of the channels • Propose functions for specific regions of the channel proteins
Amino Acid Sequence Enables Ion Channel Structure Determination
The Voltage Sensor of Ion Channels • Kv channel: Voltage sensor: S4 – alternating arg and lys residues (+). • P(O) determines the overall channel activity. • P(O) increases as the transmembraneψ is depolarized. • P(O) decreases zero at hyperpolarizing potential. • Methods used to study: - Substcys accessibility method (SCAM) – substcys at a specific location in the S4 conformational change. - FRET – shown that S4 undergoes ~180° rotation upon depolarization.
Voltage-Gated Na Channels • H5 loops form part of the ion-conducting pore. • A specific glu residue within these structures forms the binding sites for tetrodotoxin and saxitoxin. • These negatively charged residues also impart ion selectivity. • Na channels inactivate within 1-2 msec determines AP duration. • Ile-Phe-Met residues in the 3rdcytoplasmic loop (3rd-4th domain) cause the fast inactivation.
Na2+ Na2+ β1 β2 α α Out In S S
Brain Na channel = 1 pore-forming α subunit + 2 auxillaryβ subunits. • The β1 subunit responsible for fast inactivation. • The β1 and β3 isoformsmay regulate Na channel targeting to the Nodes of Ranvier. • Heart and skeletal muscle express Nav1.4 and Nav1.5 channels, respectively, where contribute to saltatory conduction of APs. • Na channels are held in place by ankyrin G complexing with NCAMs and ECM components (e.g., tenascins, phosphacans).
Voltage-Gated K Channels • Topology and structure of K channel similar to that of Na channel. • Most diverse class of ion channels. • May exist as homomers or as heteropolymers, which may exist as, e.g., A-B-A-B or A-A-B-B.
Xenopusoocytes as a Heterologous Expression system for Studying Cloned Ion Channels • Current-voltage relationship of Kv1.1 currents. • Plot the normalized peak tail currents recorded at -50 mV, as a function of the pre-pulse potentials. • Shows the fit with the Boltzmann function: I = 1/[1 + e-(V-V1/2)/k] from which the half-maximal activation voltage of the channel V1/2 and the steepness of its voltage-dependence (slope factor k) are calculated.
Inactivation Mechanisms of Kv Channels • The inactivation of delayed-rectifier K channels controls neuronal firing properties and their responses to input stimuli. • 2 Principle types of inactivation: N- and C-type. • Ball-and –chain mechanism of pore occlusion. - Occluding ball = 1st 20 N-terminal aas of Shaker channels. - 4 inactivating particles have been found, although only 1 is enough to occlude the pore. very effective occluding mechanism faster inactivation than those with only 1 particle (next slide):
Schematic Diagram of the Molecular Mechanism of N-type Inactivation • Based on x-ray crystallography (next slide). • Upon membrane depolarization, the intracellular gates open and the positively charged inactivation particle blocks the channel by entering the central cavity through 1 of the 4 windows formed by the T1 domains and the T1-S1 linkers. • 4 ‘balls and chain’ are provided by the corresponding auxiliary subunits that are anchored to the T1 domains; however, only 3 are visible in this figure (both the Kv and the β subunits are not shown for clarity). • The Kvβ subunits bind NADP+ (currently not known why)
α1 K+ α1 K+ T1 Cytoplasm Tether β Inactivation Particle
Some Kv channels inactivate slowly (C-type and P-types). • During intense neural activity, the C-type inactivation of Kv channels can accumulate, modifying both the firing rate and the shape of the AP. • Involves a conformational Δ of the extracellular mouth of the pore and a constriction of the selectivity filter.
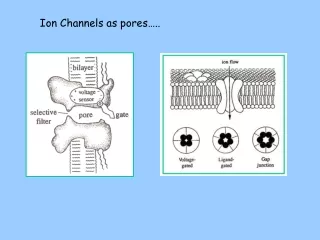
Selectivity Filter • Many channels are selective for only 1 or 2 different chemicals (ions, sugars, etc.). • The K+ channel has such a filter, which is a narrow region towards the extracellular surface of the membrane. • Two K+ ions can occupy the selectivity filter simultaneously, with a third in a H2O-filled cavity deeper in the pore.
Proposed Mechanisms for Channel Ion Selectivity Non-specific cation channel, i.e. little selectivity other than for cations 10-20 X more Na+ than K+ 100 X more K+ than Na+ Voltage-gated K+ channel – 3.3 A in diameter Voltage-gated Na+ channel - 4 A in diameter Ach receptor channel - 6.5 A in diameter
Proposed Mechanisms for Channel Ion Selectivity by Channels: Ionic size Non-specific cation channel, i.e. little selectivity other than for cations 10-20 X more Na+ than K+ 100 X more K+ than Na+ Ach receptor channel - 6.5 A in diameter Voltage-gated K+ channel – 3.3 A in diameter Voltage-gated Na+ channel - 4 A in diameter Non-hydrated Na+ ion = 1.9 A in diameter Non-hydrated K+ ion = 2.7 A in diameter If ionic size explains channel selectivity, why is the K+ channel so selective for K+ since Na+ is smaller?
Proposed Mechanisms for Ion Selectivity by Channels: Ionic size Non-specific cation channel, i.e. little selectivity other than for cations 10-20 X more Na+ than K+ 100 X more K+ than Na+ Ach receptor channel - 6.5 A in diameter Voltage-gated K+ channel – 3.3 A in diameter Voltage-gated Na+ channel - 4 A in diameter Hydrated Na+ ion = 3.3-4 A in diameter Modified Model = perhaps channels select based on hydrated ionic radius? Hydrated K+ ion = 3.3 A in diameter (K+ is larger, has a lower charge density and so attracts fewer waters of hydration.)
Proposed Mechanisms for Ion Selectivity by Channels: Ionic size The modified model explains K+ channel selectivity, i.e. the hydrated K+ just fits into the channel and the hydrated Na+ is too big to fit. However, how do we explain the +/- sodium channel selectivity? A selectivity filter exists inside the channel
Proposed Mechanisms for Ion Selectivity by Channels: Ionic size How might it work? Similar to enzymes, but much faster? Sodium recognition site = selectivity filter Na+ Na+
Evidence for a Selectivity Filter If channels are simple resistors, than movement through an open channel should be a function of the concentration gradient for the ion across the membrane Rate of ion movement = ax[ion]I/[ion]o (current flow = diffusion) Linear relationship with slope = a
Evidence for a Selectivity Filter Observed data for Na+ channel Unitary current (pa) = recordings from single channels Expected data External [Na+] mM
Evidence for a Selectivity Filter Data for voltage-gated Na+ channel do not fit the model of a channel as a simple resistor in the membrane. Instead, the current flow through the Na+ channel plateaus or “saturates” at high [Na+]. This relationship looks like what happens to an enzyme at high [substrate]. Perhaps some channels select ions based on the same biochemical mechanisms used by enzymes to select their substrates? In the end, the final determinations of channel gating mechanisms and ion selectivities will come from X-ray crystallography of the purified channels.
Inward Rectifying K Channels and Cell Excitability • 2 transmembrane domains separated by a K+ pore sequence and can assemble as both homotetramers and heterotetramers. • Acts as a diode: Iinward > Ψ< EK. But at more positive Ψs, Ioutward is inhibited and the Ψ is therefore, free to change. • Rectifying nature of the conductance is because of a voltage-dependent block of the intracellular side of the pore by cytoplasmic polyamines and Mg2+ ions.
K Selectivity of Kv Channels(next slide) • The H5 loop contributes to the ion-conducting pore. • Site-directed mutagenesis of the H5 loop: GYG. • GYG acts as the K+ selectivity filter, which is lined with carbonyl O atoms. • X-ray crystallography: 3.2A resolution; central cavity diameter = 10A, channel length = 12A. • The pore contains 2 K+ ions, 7.5A apart. • These dimensions optimal for rapid conduction and selectivity.
KcsA channel expressed in S lividans, highly homologous to the mammalian Kir channel.
Inhibitory neurotransmitters exert their inhibitory actions by activating G-protein coupled inward rectifiers. • These channels belong to the Kir3.x family. • Regulate excitability in brain and heart. • The dissociated βγ subunits stimulate heteromeric Kir3.1/Kir3.4 channel activity by physically interacting with their intracellular termini. • Recall: activation of these channels results in an efflux of K+ ions that causes membrane hyperpolarization and cell inhibition.
KATP channels – another important group of inward rectifier K channels. • Insulin secretion from pancreatic β-cells is mediated by the closure of these channels caused by increased levels of ATPcytoplasm. • KATP channel = 4 Kir6.2 subunits + 4 sulfonylurea receptors (sensitivity to drugs). • ATP inhibits channel opening by interacting with the Kir6.2 subunits. • Expressed in brain and heart, where they couple metabolic state of the cell to electrical activity.
Voltage-Gated Ca Channels • Activity initiated by depolarizing stimuli. • Ca2+ influx down a steep electrical and chemical gradient. • Can be depolarized by weak or strong stimuli: called low-threshold or high-threshold Ca2+ channels. • More detailed electrophysiological studies revealed several channel types: L-, P-, Q-, N-, R-, and T-types
VG Ca Channel Types and Voltage-Dependency • Cav1.1 L-Type High-Threshold (>-10 mV) • Cav1.2 • Cav1.3 • Cav1.4 • Cav2.1 P/Q-Type High-Threshold (>-20 mV) • Cav2.2 N-Type • Cav2.3 R-Type • Cav3.1 T-Type Low-Threshold (>-70 mV) • Cav3.2 • Cav3.3
L-Type • High To (> -10 mV). • Long-lasting openings, slow inactivation. • In muscle, are voltage sensors for excitation-contraction coupling. • In heart, β-adrenergic receptors stimulate activity of Cav1.2 channels to enhance cardiac contractility and excitability (pace maker cells). • Cav1.2 and Cav1.3 widely expressed and involved in hippocampal-dependent plasticity via NMDA receptors
P/Q-Type: • Cav2.1 (P/Q type) abundantly and widely expressed throughout the CNS, regulate fast synaptic transmisssion neuron survival, excitability, gene expression, and plasticity. • Located at the NMJ and at most presynaptic terminals in the cerebellum. • SNARE protein complex. • Undergos alt. splicing of the α1A gene. • KO mice for P/Q type exhibit ataxia, dystonia, and Purkinje cell death.
N-Type: • Expressed in sympathetic nervous system. • Voltage dependence modulated by neurotransmitters via GPCRs. • Activation of the Gβγ subunits inhibits Cav2.2 channels by decrease the mobility of the voltage sensor. • Thus, the currents show slower time courses of activation and decreased voltage-dependence.
T-Type: • Cav3.x – low threshold, characterized by transient kinetics, small single channel conductance and fast inactivation. • Various isoforms (splice variants) display various kinetics of inactivation. • Kinetics determined by the N-terminal domains of the β subunits.
Ca2+ α2 α1 S S Out γ δ In β