Download
1 / 23
290 likes | 662 Vues
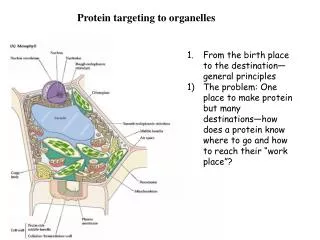
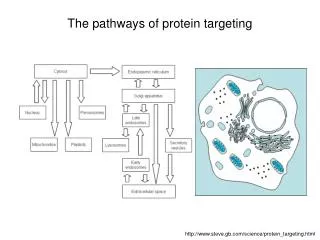
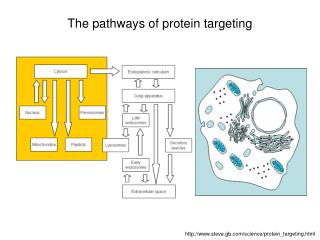
The pathways of protein targeting. http://www.steve.gb.com/science/protein_targeting.html. Mitochondria. Mitochondria have two soluble compartments: Intermembrane space Matrix ... and two membranes : Outer membrane Inner membrane (cristae)

E N D
The pathways of protein targeting http://www.steve.gb.com/science/protein_targeting.html
Mitochondria Mitochondria have two soluble compartments: • Intermembrane space • Matrix ... and two membranes: • Outer membrane • Inner membrane (cristae) • Mitochondria have their own genome and their own protein synthesizing machinery. However, not all proteins necessary for mitochondrial function are encoded by the mitochondrial genome. Most mitochondrial proteins are coded by genes in the cell nucleus and imported into the mitochondrion. The exact number of genes encoded by the nucleus and the mitochondrial genome differs between species.
Mitochondrial genome • The human mitochondrial genome is a circular DNA molecule of about 16 kilobases. It encodes 37 genes: 13 for subunits of respiratory complexes I, III, IV, and V, 22 for mitochondrial tRNA, and 2 for rRNA. One mitochondrion can contain two to ten copies of its DNA.
Import of proteins into the mitochondrion • Import of proteins into the mitochondrion is mediated by the TIM and TOM translocators, which span the inner and outer membranes, respectively. • There is no strict consensus sequence of signal peptides for the import of mitochondrial proteins. The signal is an amphipathic N-terminal α-helix with a positive stripe down one side, and a hydrophobic stripe down the other. An α-helix turns once every 3.6 amino acids, hence the approximate spacing of the blocks below • H3N+--- MLSLRQSIRFFKPAT RTL--…---COO−Part of mitochondrial import sequence from cytochrome oxidase. Note the hydrophobic phenylalanine (hexagonal) residues mostly point towards the bottom of the diagram, and the positively charged lysine and arginine residues mostly point towards the top of the diagram
The mitochondrial protein import machinery (I) • Preproteins with an amino-terminal presequence (red) as well as preproteins with internal targeting signals (blue) are recognized by receptors (R) and translocated by the general import pore (GIP) of the translocase of the outer membrane (TOM). Preproteins with a presequence are translocated across the inner membrane by the TIM23 complex. This requires the membrane potential (Δψ) and the ATP-dependent action of mitochondrial heat-shock protein 70 (mtHsp70). • The presequences are cleaved off by the mitochondrial processing peptidase (MPP) in the matrix. Preproteins with internal signals are guided by tiny Tim proteins across the intermembrane space to the TIM22 complex of the inner membrane and inserted into the membrane in a Δψ -dependent step. Pfanner and Geissler, Nature Reviews Molecular Cell Biology 2, 339-349 (May 2001)
The mitochondrial protein import machinery (II) • Variations on the two main import pathways. • Pathway A is used by presequence-carrying preproteins destined for the matrix and various preproteins destined for the outer membrane, intermem-brane space or inner membrane. Sorting signals in the latter preproteins direct their divergence from the main pathway. • Pathway B is used by the precursors of inner membrane carriers and additional inner membrane proteins with multiple internal signals. • Crossing over of import pathways • Pathway C for the precursors of the tiny Tim proteins of the intermembrane space is simple, and does not require surface receptors, but only Tom5 and Tom40 of the GIP. • The pathways D (precursor of Tim22) and E (precursor of Tim54) combine distinct portions of the two main import pathways A and B, leading to a crossing over at the level of the GIP. Pfanner and Geissler, Nature Reviews Molecular Cell Biology 2, 339-349 (May 2001)
Mitochondrial targeting sequences again Pfanner and Geissler, Nature Reviews Molecular Cell Biology 2, 339-349 (May 2001)
Mitochondrial protein import machinery (Summary) • The transport of nuclear-encoded proteins into and across the inner mitochondrial membrane is mechanistically independent of the translocation across the outer membrane. The membrane potential is only necessary for the translocation across the inner membrane. Two major translocation pathways into the inner membrane can be distinguished. • Preproteins with an amino-terminal signal sequence (presequence) are translocated by the TIM23 complex (components Tim17, Tim23 and Tim44). Tim23 forms a specific, voltage-activated channel for preproteins. • Hydrophobic proteins with multiple internal targeting signals use the same GIP of the outer membrane, but a different translocase of the inner membrane, termed the TIM22 complex. Tiny Tim proteins in the intermembrane space guide the hydrophobic proteins across this aqueous compartment to the membrane-embedded TIM22 complex (components Tim18, Tim22 and Tim54).
Chloroplasts Chloroplasts have three soluble compart-ments: • Intermembrane space • Stroma • Lumen space ...and three membranes: • Outer membrane • Inner membrane • Thylakoid membranes Similar to mitochondria, chloroplast have their own genome. As an example, the genome of the chloroplasts found in Marchantia polymorpha contains 121,024 base pairs in a closed circle. These make up some 128 genes. All gene products are used within the chloroplast, but all the chloroplast structures also depend on proteins • encoded by nuclear genes • translated in the cytosol, and • imported into the chloroplast.
Protein import into chloroplasts • Protein import into chloroplasts is much the same as in mitochondria. Hydrophobic/ positive stripes along a helix acts as signal peptides, although these must be distinguishable from mitochondrial signal, since if plastid genes are manipulated to have a mitochondrial import signal, they end up in mitochondria, rather than chloroplasts. However, there are two main differences: • GTP is required (because there is no PMF, proton motive force, across the inner membrane), and • the third compartment adds extra complication. • Lumenal translocation requires a second signal and can use four different pathways: • Sec - homologous to the translocation across a bacterial plasmalemma (the thylakoids are pinched-off vesicles from the inner membrane, which are homologous to the plasmalemma of their cyanobacterial ancestors). • SRP pathway - like RER. • Spontaneous insertion of the protein into the membrane. • ΔpH dependent, exploiting the PMF, proton motive force. across the thylakoid membrane. Soll & Schleiff, Nature Reviews Molecular Cell Biology 5, 198-208 (March 2004)
Translocon of the chloroplast envelope • The translocation of precursor proteins across the envelope membranes depends on chaperones (yellow) or processing peptidases (pale yellow), proteins in the outer envelope membrane (orange), proteins in the inner envelope membrane (blue) and stromal modulators (green). • The complex localized to the outer membrane can be dissected into a ‘core complex’ composed of the two GTPases Toc159 and Toc34, which are proposed to be regulated by phosphorylation in vitro and the translocation channel Toc75. It is proposed that Toc34 recognizes the 14-3-3 protein participating in the formation of the cytosolic guidance complex but that it also recognizes precursor proteins. Toc64 and Toc12 form a complex that is involved in perception of Hsp90-delivered precursor proteins and, together with Tic22 and imsHsp70 in the assembly of an intermembrane space (IMS) complex for the transfer across the intermembrane space. • In the inner membrane, Tic110 or/and Tic20 form the translocation pore, which together with Tic40 and the stromal stHsp93 form the translocating unit. We describe a model whereby the iron–sulfur cluster containing Tic55, the NAD(P)H and ferredoxin–NADP–oxidoreductase (FNR) binding Tic62 and the NADP(H) and calmodulin (CAL) binding Tic32 form a Tic110 associated complex regulating the activity of the translocator. The processing peptidase SPP cleaves the transit peptides in the stroma, whereas Plsp1 cleaves the transit peptide of Toc75 in the intermembrane space. TCB 18, 2008, Pages 19-27
Model for Chloroplast Protein Import Biochim. Biophys. Acta, 2001, 1541: 64-97
Model for Chloroplast Protein Import • As chloroplast preproteins emerge from 80S ribosomes, they are bound by a cytosolic guidance complex which docks at Toc64 (stage 0); certain preproteins may bypass this guidance step and proceed directly to the core Toc complex (stage 1). Preproteins unload from the guidance complex and pass to a trimeric receptor complex comprising Toc159, Toc34 and Toc75, either directly or via the lipid matrix of the outer envelope membrane. Subsequently, the import mechanism can be divided into three distinct stages, as indicated, based on energetic requirements. At stage 1 (energy-independent binding) the preprotein interacts reversibly with the heterotrimeric Toc receptor complex. Progression to stage 2 (or formation of the early import intermediate) requires ATP at low concentrations in the intermembrane space, and GTP. At this stage, the preprotein is inserted across the outer envelope membrane and is in contact with components of the Tic apparatus. Stage 3 (complete translocation) requires high concentrations of ATP in the stroma. The preprotein is translocated simultaneously across both envelope membranes at a contact site, the transit peptide is cleaved by the stromal processing peptidase (SPP) and the mature protein takes on its final conformation. Data from different laboratories suggest there may be two distinct Tic complexes (Tic complexes A and B). Tic complex B was arbitrarily chosen for the illustrated import reaction. A very similar diagram could be drawn to show an import reaction involving Tic complex A. Numbers indicate the predicted molecular weights of the proteins, and therefore identify the different Toc/Tic components. OM denotes outer envelope membrane, and IM denotes inner envelope membrane. Taken from Biochim. Biophys. Acta, 2001, 1541: 64-97.
Proteins are selectively imported into peroxisomes. Since the organelles contain no DNA or ribosomes and thus have no means of producing proteins, all of their proteins must be imported across the membrane. It is believed that necessary proteins enter through the endoplasmic reticulum during biogenesis as well as through membrane proteins.
Peroxisomal protein targeting • A specific protein signal (PTS or peroxisomal targeting signal) of three amino acids (SKL; serine-lysine-leucine) at the C-terminus of many peroxisomal proteins signals the membrane of the peroxisome to import them into the organelle. Other peroxisomal proteins contain a signal at the N-terminus. There are at least 32 known peroxisomal proteins, called peroxins, which participate in the process of importing proteins by means of ATP hydrolysis. • Proteins do not have to unfold to be imported into the peroxisome. The protein receptors, the peroxins PEX5 and PEX7, accompany their cargoes (containing a PTS1 or a PTS2, respectively) all the way into the peroxisome where they release the cargo and then return to the cytosol - a step named recycling. • Overall, the import cycle is referred to as the extended shuttle mechanism. Evidence now indicates that ATP hydrolysis is required for the recycling of receptors to the cytosol. Also, ubiquitination appears to be crucial for the export of PEX5 from the peroxisome, to the cytosol. Little is known about the import of PEX7, although it has helper proteins that have been shown to be ubiquitinated.
Protein import into peroxisomes • The peroxisomal import receptors peroxin-5 (Pex5) and Pex7 bind to specific peroxisome targeting signals (PTSs) in peroxisomal matrix proteins in the cytosol and direct these proteins to the peroxisomal membrane. Most peroxisomal matrix proteins carry a tripeptidePTS1 at their extreme C terminus, which has the consensus sequence (S/A/C)-(K/R/H)-L and is recognized by Pex5. PTS2 is an N-terminal nonapeptide, which has the consensus motif (R/K)-(L/I/V)-X5-(H/Q)-(L/A/F) (X is any amino acid) and is recognized by Pex7. • The cycling of receptors can be conceptually divided into four steps (see figure, part a, which shows the steps for Pex5): first, the assembly of the receptor–cargo complex in the cytosol (recognition); second, binding of the receptor–cargo complex at the peroxisomal membrane (docking); third, the release of cargo (dissociation); and fourth, receptor export to the cytosol to allow for another round of import (recycling). There are different views regarding where the dissociation of the receptor–cargo complex occurs (part b). The cargo might be released on the cytosolic side of the membrane (the 'simple shuttle model'), on the matrix side of the membrane (membrane shuttle: the 'trap door model', the 'membrane-associated cargo release model' or the 'transient pore model' discussed in this article), or after the complete translocation of the receptor–cargo complex into the matrix of the peroxisome (the 'extended shuttle model').
Peroxin-7 • Peroxin-7 Pex7p) is a receptor associated with Refsum's disease and rhizomelic chondrodysplasia punctata type 1. Pex7p accompanies cargo proteins with the PTS2 signal to their destination in the peroxisome.
Nucleus • For most macromolecules, getting in and out of the nucleus is hampered by the double membrane that surrounds it. Some small, nonpolar molecules can diffuse through the membranes (particularly important here are the steroid hormones), but much else goes via the nuclear pores. • Nucleus → cytoplasm: mRNA, ribosomes. • Cytoplasm → nucleus: proteins (DNA polymerase, lamins), lipids, carbo-hydrates, signalling mole-cules.
Nuclear pore complex • The nuclear pores, which are composed of (the imaginatively named) nucleoporin proteins, have an 8-fold symmetry, with an outer diameter of c. 100 nm, and an inner diameter in electron micrographs of c. 80 nm. The pore itself is only c. 9 nm (free diffusion), but dilates to 30 nm (active transport). Some nucleoporins (particularly the central 'plug') are sometimes lost during preparation. The cytoplasmic ring of the pore has fibrils, which are though to form queues of chemicals entering the nucleus. The nucleoplasmic ring has an attached 'basket'. Note the way the luminal nucleoporins clamp the doubled-back nuclear envelope. • Nuclear pore from above (left) and from the side (right) http://mol-biol4masters.org/Co_and_Post_Translational_Events5-Cellular_Protein_Traffic.htm
Nuclear pore • The nuclear pore complex is a huge structure with hundreds of proteins; the molecular weight of the total mass is 125 million Daltons or more. The pore complex is constructed like an annular concentric rings looks like a wine barrel with an upper ring and lower ring of eight subunits each. The upper top ring faces the cytoplasm and lower or bottom ring faces the nuclear sap. Protein filaments emanate from the upper ring, so also from the bottom ring, but the filaments from the bottom ring are connected to each other in the form of a ring. It contains a central channel of 40 nm size runs all along the length of the barrel. The central channel is linked to outer coaxial ring of proteins by eight spokes. The coaxial ring in turn is connected to nuclear membranes by radial arms. The space between the radial spokes also provides free space for the movement of smaller molecules. The central pore complex has cytoplasmic Iris and nucleoplasmic Iris connected to cytoplasmic ring and nucleoplasmic ring, respectively. • A network of structural proteins (Lamins) supports the nuclear membrane at its inner surface. The number of pore complexes can expand and contract according to the needs. Average number of pore complexes per nucleus is ~3000. • The pore complex is responsible for the import of several kinds of nuclear proteins and export of several species of RNAs, ribosomal subunits and many snRNPs. The number of proteins imported per pore complex per minute is remarkable. One hundred Histones are imported per minute per pore complex. Non-histones 200, and riboproteins 150 per minute imported. The same pore complexes export ~5 ribosomes per minute per pore complex, mRNAs one per minute. The database estimates 6000 to 10000 different proteins are imported from cytoplasm into the nucleus. • Molecules of 5 to 500 Daltons size easily move across the pore complex, thus nucleotides, ions many other molecules have easy access into the nucleus. But proteins of more than 50KD (size of ~5nm) cannot diffuse passively, but they are transported in ATP dependent manner (active process). The central channel is exclusively used for the transport larger cargo by expansion and contraction mode. They can transport gold particles of size~ 20nm through central core. The pore complex transports ribosome of 120 x 200Å size. The central channel has a cytoplasmic iris connecting cytoplasm and a nucleoplasmic iris connecting nucleoplasm. Though the pore complex allows components passive diffusion, it is always facilitated. Active transport is always a facilated transport. Cytoplasm contains 20-30 thousand different proteins, but only those proteins that are required are transported and not others. So also export is directed and facilitated. http://mol-biol4masters.org/Co_and_Post_Translational_Events5-Cellular_Protein_Traffic.htm
Import of nuclear proteins • Transport of proteins into the nucleus is determined by specific signal sequences called Nuclear Localization Sequences (NLS). Most of the nuclear proteins have a common signature sequences for the import. Similarly those proteins or RNA species destined for export should have similar signature sequences called Nuclear Export Sequences (NES). Proteins involved in import are called Importins and those used in export Exportins. • Proteins to be imported into the nucleus have a nuclear import peptidePPKKKRKV (proline, lysine, arginine, valine) - the import signal binds FG-repeats in nuclear-import receptors (karyopherins, also known as importins). Export protein sequences have a conserved 10 amino acids with conserved leucines. • Import of nuclear protein requires a host of proteins called Importins. But the importin-α, β, and β3 are important. The importins are located at the orifice of the pore complex. Importin-α binds to NLS sequences then transfers to importin-β, which in turn delivers to nucleoporins (Nuc-Ps) found at the nuclear membrane in the pore complex. They are involved in inward movement of cargo. There are a many nucleoporins involved in translocation of proteins into the nucleus, it requires the input of ATP energy. There is another group of importins called importin-β3, another group is called Transportins. which not only binds to NLS sequences but also delivers the cargo to nucleoporins. Transportin M9 is mainly responsible for the transport of hnRNPs. Importinβ3 is belied to responsible for the import of riboproteins. There are more importins, which carry signal molecules into the nucleus for activating transcription. Nucleoporins have repeated motifs such as GKFG, FG, and FxFG. These motifs help in binding to importin-β and driving the cargo inside. • Another cytosolic factor that supports translocation is Ran. It is a GTPase protein. Ran is a monomeric G-protein. The Ran-GTP complex is found associated with importins at cytosolic side and also found at the nucleoplasmic side. The Ran-GTP is active, when hydrolysed it provides energy for the movement. The Ran-GDP, is inactive, it will be activated only when GDP is exchanged with another GTP by another factor called GEF (GTP exchange factor). http://mol-biol4masters.org/Co_and_Post_Translational_ Events5-Cellular_Protein_Traffic.htm
Nuclear export • Similar to importins there are Exportins molecules that are specific each type and they are responsible for the export of mRNAs, ScRNAs, SnRNAs tRNAs, many proteins and ribosomes into cytoplasm. Export of protein requires signal sequences such as NES (nuclear export sequence), a ten amino acid residues stretch that is rich in Leucines. Export of components is species specific and requires specific nucleoporins aided by Ran-GTP factors found at nucleoplasmic surface. http://mol-biol4masters.org/Co_and_Post_Translational_Events5-Cellular_Protein_Traffic.htm
Nuclear RNA export Köhler & Hurt, Nature Reviews Molecular Cell Biology 8, 761-773 (2007) • The major RNA export routes are shown (tRNA, microRNA (miRNA), small nuclear (sn)RNA, mRNA, ribosomal (r)RNA). In each case, the primary RNA transcript is shown, as well as the transport-competent RNA after it has undergone processing, maturation and assembly with export factors (export adaptors are shown in blue, export receptors are shown in yellow). Prominent structural motifs in pre-RNAs are indicated (single/double-stranded RNA, loops, exons and introns, 5' cap and 3' poly(A) tail). For the mRNA export route, the names of both metazoan and yeast proteins are indicated, and mRNAs are shown with additional adaptor proteins and RNA-binding factors (orange ovals). In the case of rRNA, the general exporter in eukaryotes, CRM1, and two auxiliary exporters, Mex67–Mtr2 and Arx1, that have only been studied in yeast are depicted. CBC, cap-binding complex; Exp, exportin.