Download
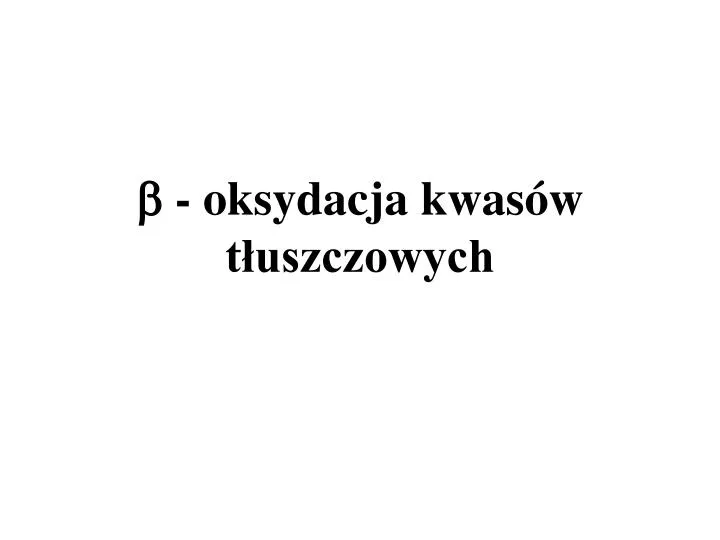
1 / 30
390 likes | 1.38k Vues
- oksydacja kwasów tłuszczowych. Nazwa przemiany: -oksydacja , pochodzi od nazwy trzeciego węgla w łańcuchu kwasu tłuszczowego. Węgiel w pozycji jest najwyżej utlenionym podczas redoksowych reakcji katabolicznych kwasów tłuszczowych.

E N D
Nazwa przemiany: -oksydacja, pochodzi od nazwy trzeciego węgla w łańcuchu kwasu tłuszczowego. Węgiel w pozycjijest najwyżej utlenionym podczas redoksowych reakcji katabolicznych kwasów tłuszczowych. Acylo CoAjest wtedy całkowicie rozkładany do kwasu octowego (w postaci acetylo CoA.) Proces -oksydacji zachodzi w matriks mitochondrialnej.
Mobilizacja kwasów tłuszczowych w adipocytach jest indukowana hormonalnie.
Aktywacja i transport kwasów tłuszczowych do mitochondriów Acylotransferaza karnitynowa Translokaza acylokarnitynowa
Karnityna Nazwa systematyczna: kwas3-hydroksy-4-(trimetylo-amonio)butanowy Karnityna jest czwartorzędową zasadą amoniową tworzoną z aminokwasów lizynyi metioniny. W komórkach służy jako przenośnik kwasów tłuszczowych z cytozolu do mitochondriów, podczas rozkładu tłuszczów w celach energetycznych. Jest w sprzedaży jako suplement diety.
Obok: cztery podstawowe reakcje -oksydacji w matrix mitochondrialnej. Acetylo-CoA – główny produkt, jest katabolizowany w Cyklu Krebsa do CO2 i H2O.
Oksydacja nienasyconych kwasów tłuszczowych Utlenianie nienasyconych kwasów tłuszczowych biegnie w zasadziew ten sam sposób jak nasyconych;wyjątkiem są te fragmenty, które dotyczą wiązań podwójnych.Wiązania takie podlegają izomeryzacji przy udziale specyficznejisomerazyenoilo-CoA, po czym utlenianie jest kontynuowane. W przypadku linolenianu, obecność dodatkowego wiązania nienasyconego(Δ12) powoduje powstanie Δ2,4dienoilo-CoA. Taka cząsteczka wymaga dodatkowego enzymu, reduktazyNADPH 2,4-dienoilowej, koniecznej do likwidacji dalszego wiązania podwójnego (Δ4)
Beta oksydacja kwasów o nieparzystej liczbie wiązań podwójnych Kwasy tego typu powszechnie występują u roślin i organizmów morskich.Zatem ssaki, które spożywają taki pokarm muszą mieć możliwość metabolizowania nieparzystych kwasów w procecie beta oksydacji. Wtedy końcowym produktem przemiany jest trójwęglowa cząsteczka propionylo-CoAzamiast acetylo-CoA. Propionylo-CoA jest także produktem przemian izoleucyny, waliny i metioniny. Aby wejść do cyklu Krebsa propionylo-CoA musi ulec przemianie do bursztynylo-CoA.
Regulacja -oksydacji FFA Malonylo-CoAspowalnia wprowadzanie reszt acylowych do mitochondriów poprzez hamowanieodpowiedzialnej za ten transportacylotranferazykarnitynowej. Spowalnia zatem również beta oksydację. Gdy rośnie poziom acylo-CoA, beta oksydacja jest stymulowana. Jednak podwyższony poziomcytrynianuhamuje beta oksydację, ponieważ oznacza onrównież wysokie stężenie acetylo-CoA. Sam acetylo-CoA także bezpośrednio jestinhibitorem beta oksydacji na poziomie reakcji tiolizy.Zatem cały system działa na zasadzie typowego, ujemnego sprzężenia zwrotnego.
Ketogeneza Podczas wysokich obrotów spirali reakcji beta oksydacji, może dojść do powstania olbrzymich ilości acetylo-CoA. Dotyczy to szczególnie wątroby. Gdy dojdzie tam do przekroczenia możliwości katabolicznych cyklu cytrynianowego, uruchomiony zostaje proces ketogenezy. Powstają wtedy tzw ciała ketonowe:acetooctan, β-hydroksymaślaniaceton.
HMG-CoA jest „strategicznym” metabolitem nie tylko w -oksydacji ale również procesie syntezy steroidów w cytosolu.
Ciała ketonowe są ważnym „paliwem” dla tkanek poza-wątrobowych. Acetooctan jest tam aktywowany przez specyficzną transferazę, która przenosi koenzym A z burszynylo-CoA. Acetoacetoacetylo-CoA jest rozczepiany przez tiolazę do acetylo-CoA i utleniany w cyklu Krebsa. Ten typ „paliwa” jest szczególnie istotny dla komórek mózgu oraz mięśnia sercowego.
Patologiczne aspekty ketogenezy Niskie stężenia ciał ketonowych zdarzają się w warunkach fizjologicznych, natomiast ich długotrwały wysoki poziom jest oznaką zaburzeń metabolizmu. Ketonemia (ciała ketonowe we krwi) i ketonuria (ciała ketonowe w moczu) są objawem cukrzycy, głodu, lub kwasicy alkoholików. Przeżuwacze mają fizjologicznie wyższe stężenie ciał ketonowych we krwi i w moczu i z tego powodu są bardziej narażone na kwasicę ketozową niż inne grupy zwierząt. Każde zakłócenie procesu trawienia spowodowane błędami dietetycznymi może spowodować stan patologiczny. Jest to szczególnie ważne u wysoko-mlecznych krów, u których niemal cała pula glukozy wyprodukowanej w wątrobie z kwasu propionowego, zostaje zużyta do syntezy laktozy w gruczole mlekowym. „Tłuszcze spalają się w płomieniu węglowodanowym” Slogan ten oznacza, że każda dodatkowa cząsteczka acetylo-CoA musi znaleźć dodatkową cząsteczkę szczawiooctanu (powstałą z pirogronianu), aby wejść do cyklu cytrynianowego. W przeciwnym przypadku pojawiają się ciała ketonowe.
ProdukcjaATP podczas -oksydacji palmitoilo-CoA Kwas palmitynowy (heksadekanowy) posiada łańcuch o16 atomach węgla, wymaga więc 7 cykli oksydacyjnych do produkcji 8 cząsteczek acetylo-CoA: 1-sze utlenienie: 7 FADH2x 1.5 ATP= 10.5 ATP 2-gie utlenienie: 7NADHx 2.5 ATP= 17.5 ATP Cykl Krebsa:8 Acetylo CoAx10 ATP= 80.0 ATP Aktywacja = -2.0 ATP Netto = 106 ATP
Synteza kwasów tłuszczowych polega na ich formowaniu z acetylo-CoAimalonylo-CoAprzy udziale wieloenzymatycznego kempleksu zwanego syntazą kwasów tłuszczowych.
Wstępna synteza malonylo-CoAHCO3- + ATP + acetylo-CoA ADP + Pi + malonylo-CoA KarboksylazaAcetylo-CoA katalizuje dwuetapową reakcję, w której cząstka acetylo-CoA jest karboksylowana do malonylo-CoA. Grupą prostetyczną tego enzymu jestbiotyna. Jest ona karboksylowana z udziałem ATP w jednym z miejsc aktywnych enzymu (1) a następnie przenoszona do miejsca wiązania acetylo-CoA (2), gdzie zachodzi właściwa reakcja karboksylacji.
Synteza kwasów tłuszczowych U bakteriijest katalizowana przez sześć różnych enzymów oraz białko przenoszące acyl (Acyl Carrier Protein – ACP). Ussakówjest katalizowana przez indywidualne domenybardzo dużego polipeptydu zawierającego również element przenoszący acyl.Ewolucja syntazy ssaków polegała więc na fuzji genów wszystkich białek biorących udział w tym procesie. U eukariontów kwasy tłuszczowe są syntetyzowane w cytozolu (do długości 16 atomów węgla), a następnie wydłużane (elongacja) w mitochondriach lub w siateczce ER. Desaturacja kwasów tłuszczowych (tworzenie wiązań podwójnych) wraz z elongacją nienasyconych kwasów zachodzi na błonach retikulum endoplazmatycznego.
Sekwencja reakcji podczas syntezy kwasów tłuszczowych jest dokładnym odwróceniem -oksydacji. Proces jest jednak katalizowany przez inne enzymy oraz w innym środowisku.
Białko Przenoszące AcylAcyl Carrier Protein (ACP)Grupą prostetyczną tego białka jest fosfopantoteina. Jest to ten sam element strukturalny, który występuje w koenzymie A. Tutaj pełni funkcję elastycznego ramienia przenoszącego substrat do kolejnych centrów aktywnych. Jego elementem wiążącym acyl jest grupa SH
Nazwy „katalitycznych aktywności” syntazy • Syntaza -ketoacyl – ACP • Transacetylaza malonylo CoA – ACP 3. Transacetylaza acetylo CoA – ACP 4. Dehydrataza -hydroxyacyl – ACP 5. Reduktaza enoilo-ACP 6. Reduktaza -ketoacylowa 7. Tioesteraza palmitoilowa
Sekwencja poszczególnych reakcji syntezy kwasów tłuszczowych Reszta acetylowa jest wymagana jako”primer”, natomiast trójwęglowe resztymalonylowe są donorami reszt dwuwęglowych do wydłużania nasyconego łańcucha kwasu umocowanego na ACP.Kwas ten w postaci cząsteczek acylo-ACP (W przypadku podanym obok buturylo -ACP), posłuży jako primer w następnych cyklach syntezy. NADPH jest natomiast donorem wodorów zarówno dla redukcji reszt ketonowychjak i enoilowych.
Elongacja kwasów tłuszczowych U eukariontów, produkcja nasyconych, długołańcuchowych kwasów tłuszczowych (C20 – C26) orazkwasów wielo- nienasyconych, przebiega niezależnie od FAS, przy udziale enzymów związanych z cytozolową stroną błon retikulum endoplazmatycznego (ER). Większość eukariontów posiada systemy elongacyjne uformowane w postać kompleksów wieloenzymatycznych. Mitochondria również mają pewną możliwość wydłużania kwasów tłuszczowych powyżej 16 atomów węgla. Bardzo długie kwasy tłuszczowe są używane jako prekursory w syntezie sfingolipidów, tworzących miejsca kontaktu z białkami pośredniczącymi w transdukcji sygnałów przez błony komórkowe.
Desaturacja kwasów tłuszczowych Desaturazy kwasów tłuszczowych są enzymami, które usuwają po dwa wodory z łańcucha kwasu tłuszczowego, tworząc wiązanie podwójne pomiędzy atomami węgla. Desaturazy dzieli się na dwie grupy: Delta (Δ) i Omega (ω). Grecka litera Δ (wraz z indeksem) wskazuje, że wiązanie podwójne powstaje w ustalonej pozycji od strony grupy karboksylowej kwasu.Np. desaturaza Δ6tworzy podwójne wiązanie za szóstym atomem węgla danego kwasu. Omega (ω) - wskazuje powstanie wiązania podwójnego bliżej końca metylowego długołańcuchowego kwasu. Pozycja liczona od węgla metylowego jest podana indeksem liczbowym (np. ω3). Ssaki nie posiadają tego typu enzymów i dlatego niektóre wielonienasycone kwasy są dla nich witaminami.
Krótka kaskada reakcji redoksowych jest zaangażowana w proces desaturacji. Przykład: Wprowadzenie podwójnego wiązania pomiędzy węglem 9-tym a 10-tym stearoilo-CoA wymaga tlenu cząsteczkowego oraz ferrocytochromu b5. Desaturazastearoilowaużywa NAD jako kofaktora pierwszej reakcji redoksowej: