Download

1 / 1
30 likes | 187 Vues
Overlap of the human CD8 + T cell receptor repertoire Harlan S. Robins 1,2 , SK Srivastava 1 , P Campregher 1 , CJ Turtle 1 , J Andriesen 2 , SR Riddell 1 , CS Carlson 1,2 , EH Warren 1 Fred Hutchinson Cancer Research Center, Seattle, WA 1 , Adaptive TCR Technologies, Seattle, WA 2.

E N D
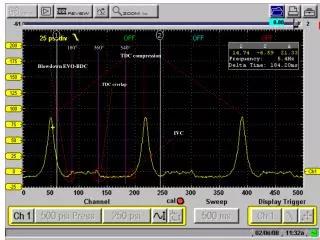
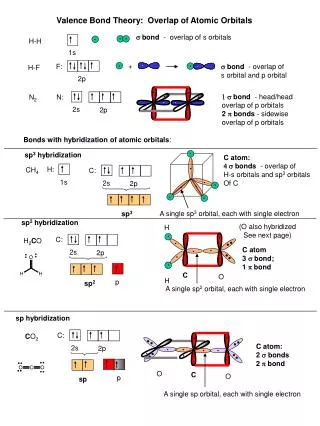
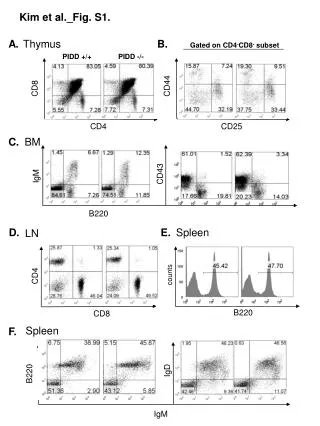
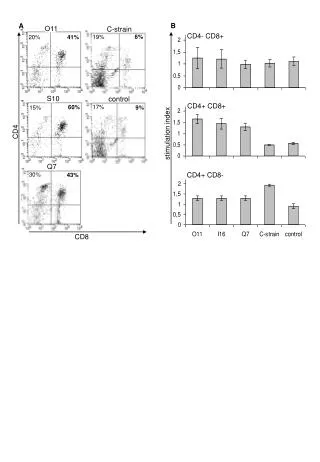
Overlap of the human CD8+ T cell receptor repertoire Harlan S. Robins1,2, SK Srivastava1, P Campregher1, CJ Turtle1, J Andriesen2, SR Riddell1, CS Carlson1,2, EH Warren1 Fred Hutchinson Cancer Research Center, Seattle, WA1, Adaptive TCR Technologies, Seattle, WA2 • Results • Millions of unique TCRB CDR3 sequences were identified (Table 2). • Table 2. Summary of the TCRB CDR3 sequence data • Utilization of specific Vβ and Jβ segments is variable within an individual, but relatively consistent between individuals (Fig. 2). • Fig. 2. Non-uniform utilization of Vβ–Jβ gene combinations in CD8+ T cells of seven healthy adults. Mean utilization frequency of specific Vβ–Jβ gene segment combinations in TCRB CDR3 sequences expressed in (A) naïve and (B) memory CD8+ T cells. • Results • Utilization of Dβ-Jβ gene segment pairs is non-random. The Dβ2-Jβ1 gene segment pair is observed less often relative to other Dβ-Jβ gene segment pairs (Fig. 3) • Fig. 3. Relative utilization of specific Dβ-Jβ gene segment pairs in naïve and memory CD8+ T cells. The proportion of all in-frame, TCRB CDR3 sequences obtained from (A) naïve or (B) memory CD 8+ T cells that utilize each Dβ-Jβ pairing. Gene pairings include a D segment, Dβ1 or Dβ2, and Jβ family, Jβ1 or Jβ2. • The actual TCRB CDR3 repertoire overlap between any two individuals is tens of thousands of amino-acid sequences (Fig. 4). • Fig 4. Calculation of the total number of TCRB CDR3 amino acid sequence overlaps between the naïve CD8+ T cell compartments of two donors. The colored points indicate the observed number of naïve and memory CD8+ TCRB CDR3 amino-acid sequences between donor 3 and the other donors. The solid lines indicate the estimated total number of CD8+ TCRB CDR3 amino-acid sequences donor 3 shares with the other donors. • Results • Unexpectedly high TCRB CDR3 amino-acid sequence overlap in both the CD8+ naïve and memory compartments (Fig. 5a). • Shared naïve and memory CD8+ T cells have fewer non-template insertions than nonshared CD8+ T cells (Fig. 5b, 5c). • Fig. 5. Characteristics of public CD8+ TCRB CDR3 amino-acid sequences. (A) Number of shared sequences in the naive and memory CD8+ CDR3 repertoires of every possible pair of individuals. A colored shape represents the relationship of a pair of individuals: HLA identical siblings (blue circle), mother daughter pair (red square), and unrelated individuals (black triangle). ( B and C) Frequency distribution of shared (red) and nonshared (blue) CDR3 sequences observed in the (B) naïve and memory (C) CD8+ compartments of every possible pair of individuals as a function of the total number of nucleotide insertions at the Vβ-Dβ and Dβ-Jβ junctions. • Conclusions • We sequenced, from the peripheral blood CD8+ T cell compartment of seven healthy adults, ~40 million rearranged TCRB CDR3 sequences. Approximately 3 million of these sequences were unique. • The realized CDR3 sequence repertoire within each individual is strongly biased toward specific Vβ–Jβ pair utilization, and dominated by sequences containing fewer than 8 non-template nucleotide insertions. • The overlap in the naïve CD8+ CDR3 sequence repertoires of any two of the individuals is ~7000 fold larger than predicted. • Introduction • The antigenic specificity of T lymphocytes is in large part determined by the amino acid sequence in the hypervariablecomplementarity-determining region 3 (CDR3) of the α and β chains of the T cell receptor (TCR). The nucleotide sequences that encode the CDR3 regions are generated by somatic rearrangement between variable (V), diversity (D), and joining (J) region gene segments. Diversity generated by the somatic rearrangement of germline genomic DNA is augmented by the deletion of nucleotides adjacent to the recombinational signal sequences (RSSs) of the V, D, J segments and template-independent insertion of nucleotides at the Vβ-Dβ, Dβ-Jβ, and Vβ-Jβ junctions. Together, these processes allow for tremendous variation within the CDR3 region. High-throughput sequencing on the immunoSEQ system enables us to globally capture the TCRB CDR3 repertoires of multiple individuals. We examine the repertoire of TCRB CDR3 sequences expressed in the naïve and memory CD8+ T cells from the peripheral blood of seven healthy adults, and assess the extant of overlap between any two individuals TCRB CDR3 repertoires. With this information we: • Estimate the utilization of Vβ–Jβ andDβ-Jβ gene segmentpairs in CD8+ T cells across seven healthy donors. • Estimate the extent to which CD8+ T cell TCRB CDR3 repertoire overlaps between any two individuals. • Screen for characteristics that distinguish shared and nonshared TCRB CDR3 sequences. • Materials and Methods • PBMC collected from seven HLA typed donors (Table 1). • Table 1: Donor characteristics • PBMC were FACS-sorted to isolate and collect memory and naïve CD8+αβ T cells. • Genomic DNA was extracted. • TCRB sequences were amplified and sequenced using the immunoSEQ assay (Fig. 1) • Fig. 1: Assay A B A A Number of shared CDR3 sequences in CD8+ naive Number of shared CDR3 sequences in CD8+ memory C B Probability of insertion B Total number of nucleotide insertions For additional information about immunoSEQ assays and the immunoSEQ Analyzer suite of bioninformatics applications at Adaptive TCR Technologies, visit our booth or contact us on the web at www.adaptivetcr.com and www.immunoseq.com. This work is published in Science Translational Medicine, September 2010, Vol 2, Issue 47. Adaptive TCR Technologies Suite 300 307 Westlake Ave N Seattle, WA 98109