Download
1 / 37
460 likes | 1.17k Vues
Chemiosmotic theory of oxidative phosphorylation. Inhibitors and uncouplers of oxidative phosphorylation. A PROTON GRADIENT POWERS THE SYNTHESIS OF ATP. The transport of electrons from NADH or FADH 2 to O 2 via the electron-transport chain is exergonic process:

E N D
Chemiosmotic theory of oxidative phosphorylation. Inhibitors and uncouplers of oxidative phosphorylation.
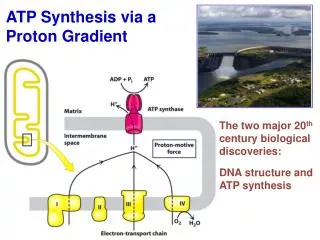
A PROTON GRADIENT POWERS THE SYNTHESIS OF ATP The transport of electrons from NADH or FADH2 to O2 via the electron-transport chain is exergonic process: NADH + ½O2 + H+ H2O + NAD+ FADH2+½O2 H2O + FAD+ Go’ = -52.6 kcal/mol for NADH -36.3 kcal/mol for FADH2 How this process is coupled to the synthesis of ATP (endergonic process)?ADP + Pi ATP + H2OGo’=+7.3 kcal/mol
The Chemiosmotic Theory • Proposed by Peter Mitchell in the 1960’s (Nobel Prize, 1978) • Chemiosmotic theory: electron transport and ATP synthesis are coupled by a proton gradient across the inner mitochondrial membrane Mitchell’s postulates for chemiosmotic theory • Intact inner mitochondrial membrane is required • Electron transport through the ETC generates a proton gradient • 3. ATP synthase catalyzes the phosphorylation of ADP in a reaction driven by movement of H+ across the inner membrane into the matrix
+ + + + + + - - - - Overview of oxidative phosphorylation As electrons flow through complexes of ETC, protons are translocated from matrix into the intermembrane space. The free energy stored in the proton concentration gradient is tapped as protons reenter the matrix via ATP synthase. As result ATP is formed from ADP and Pi.
An artificial system demonstrating the basic principle of the chemiosmotic hypothesis Synthetic vesicles contains bacteriorhodopsin and mitochondrial ATP synthase. Bacteriorhodopsin - protein that pumps protons when illuminated. When the vesicle is exposed to light, ATP is formed.
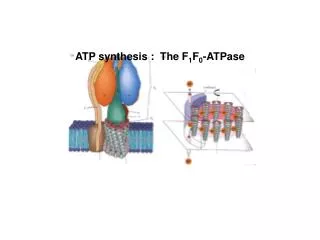
ATP Synthase Two units, Fo and F1(“knob-and-stalk”; “ball on a stick”) F1 contains the catalytic subunits where ADP and Pi are brought together for combination. F0spans the membraneand serves as a proton channel. Energy released by collapse of proton gradient is transmitted to the ATP synthesis.
F1contains 5 types of polypeptide chains -a3b3gde • Fo - a1b2c10-14(csubunits form cylindrical, membrane-bound base) • Fo and F1 are connected by agestalk and by exterior column (a1b2 andd) • The proton channel – between c ring and a subunit.
there are 3 active sites, one in eachbsubunit • c-e-gunit forms a “rotor” • a-b-d-a3b3unit is the “stator” • passage of protons through the Fo channel causes the rotor to spin • rotation of thegsubunit inside thea3b3hexamer causes domain movements in theb-subunits, opening and closing the active sites
Each b subunit contains the catalytic site. At any given time, each site is in different conformation: open (O), loose (L) or tight (T). O conformation binds ADP and Pi The affinity for ATP of T conformation is so high that it converts ADP and Pi into ATP.
Binding-Change Mechanism of ATP Synthase 1. ADP and Pi bind to an open site 2. Passage of protons causes each of three sites to change conformation. 3. The open conformation (containing the newly bound ADP and Pi) becomes a loose site. The loose site filled with ADP and Pi becomes a tight site. The ATP containing tight site becomes an open site. 4. ATP released from open site, ADP and Pi form ATP in the tight site
Experimental observation of ATP synthase rotation • Fluorescent protein arm (actin) attached togsubunits • a3b3subunits bound to a glass plate • Arm seen rotating when ATP added (observed by microscopy)
MOVEMENT ACROSS THE MITOCHONDRIAL MEMBRANES Electrons from Cytosolic NADH Enter Mitochondria by Shuttles NADH is generated in the cytosol in glycolysis. The inner mitochondrial membrane is impermeable to NADH and NAD+. Electronsfrom NADH,but not NADH itself, are carried across the mitochondrial membrane. Two shuttles move electrons: glycerol 3-phosphateshuttleandmalate-aspartateshuttle
Glycerol 3-phosphate shuttle Active in skeletal muscles and brain. Electrons enter the electron-transport chain via complex II. Therefore only 1.5 molecules of ATP are produced.
Malate-aspartate shuttle Active in heart and liver.2.5 molecules of ATP are produced.
Active Transport of ATP, ADP and Pi Across the Inner Mitochondrial Membrane • ATP must be transported to the cytosol, and ADP and Pi must enter the matrix • ADP/ATP carrier, adenine nucleotide translocase, exchanges mitochondrial ATP4- for cytosolic ADP3- • The exchange causes a net loss of -1 in the matrix (draws some energy from the H+ gradient) • Phosphate (H2PO4-) is transported into matrix in symport with H+. Phosphate carrier draws on pH. • Both transporters consume proton-motive force
Mechanism of ATP and ADP Transport ATP-ADP translocase is abundant in the inner mitochondrial membrane (about 14% of the protein) The entry of ADP into the matrix is coupled to the exit of ATP.
Mitochondrial Transporters ATP-ADP translocase – antiport of ATP and ADP Phosphate carrier – antiport of H2PO4- and OH- (symport of H2PO4- and H+) Dicarboxylate carrier – antiport of malate, succinate, or fumarate and H2PO4- Tricarboxylate carrier – antiport of citrate and H+ and malate Pyruvate carrier – antiport of pyruvate and OH- (symport of pyruvate and H+)
REGULATION OF OXIDATIVE PHOSPHORYLATION Coupling of Electron Transport with ATP Synthesis Electron transport is tightly coupled to phosphorylation. ATP can not be synthesized by oxidative phosphorylation unless there is energy from electron transport. Electrons do not flow through the electron-transport chain to O2 unless ADP is phosphorylated to ATP. Important substrates: NADH, O2, ADP Intramitochondrial ratio ATP/ADPis a control mechanism High ratio inhibits oxidative phosphorylation as ATP allosterically binds to a subunit of Complex IV
Respiratory control The most important factor in determining the rate of oxidative phosphorylation is the level of ADP. The regulation of the rate of oxidative phosphorylation by the ADP level is calledrespiratory control
Uncoupling of Electron Transport with ATP Synthesis Uncoupling of oxidative phosphorylation generates heat to maintain body temperature in hibernating animals, in newborns, and in mammals adapted to cold. Brown adipose tissues is specialized for thermogenesis. Inner mitochondrial membrane contains uncoupling protein (UCP), or thermogenin. UCP forms a pathway for the flow of protons from the cytosol to the matrix.
Uncouplers • Uncouplers are lipid-soluble aromatic weak acids • Uncouplers deplete proton gradient by transporting protons across the membrane 2,4-Dinitrophenol: an uncoupler • Because the negative charge is delocalized over the ring, both the acid and base forms of DNP are hydrophobic enough to dissolve in the membrane.
Specific inhibitors of electron transport chain and ATP-synthase Specific inhibitors of electron transport are invaluable in revealing the sequence of electron carriers. Rotenoneand amytalblock electron transfer in Complex I. Antimycin Ainterferes with electron flow thhrough Complex III. Cyanide, azide,and carbon monoxideblock electron flow in Complex IV. ATP synthaseis inhibitedbyoligomycin which prevent the influx of protons through ATP synthase.
3 4 4 2 ATP Yield Ten protons are pumped out of the matrix during the two electrons flowing from NADH to O2 (Complex I, III and IV). Six protons are pumped out of the matrix during the two electrons flowing from FADH2 to O2 (Complex III and IV). Translocation of 3H+ required by ATP synthase for eachATP produced 1 H+ needed for transport of Pi. Net:4 H+ transported for each ATP synthesized For NADH: 10 H+/ 4H+) = 2.5 ATP For FADH2: 6 H+/ 4 H+ = 1.5 ATP
Citric acid cycle supplies NADH and FADH2 to the electron transport chain Fatty Acids Acetyl Co A Amino Acids Pyruvate Glucose
Reduced coenzymes NADH and FADH2 are formed in matrix from: (1) Oxidative decarboxilation of pyruvate to acetyl CoA (2) Aerobic oxidation of acetyl CoA by the citric acid cycle (3) Oxidation of fatty acids and amino acids The NADH and FADH2 are energy-rich molecules because each contains a pair of electrons having a high transfer potential.
Electrons of NADH or FADH2 are used to reduce molecular oxygen to water. A large amount of free energy is liberated. The electrons from NADH and FADH2are not transported directly to O2 but are transferred through series of electron carriers that undergo reversible reduction and oxidation.
The flow of electrons through carriers leads to the pumping of protons out of the mitochondrial matrix. The resulting distribution of protons generates a pH gradient and a transmembrane electrical potential that creates a protonmotive force.
ATP is synthesized when protons flow back to the mitochondrial matrix through an enzyme complex ATP synthase. The oxidation of fuels and the phosphorylation of ADP are coupled by a proton gradient across the inner mitochondrial membrane. Oxidative phosphorylation is the process in which ATP is formed as a result of the transfer of electrons from NADH or FADH2 to O2 by a series of electron carriers.
OXIDATIVE PHOSPHORYLATION IN EUKARYOTES TAKES PLACE IN MITOCHONDRIA Two membranes: outer membrane inner membrane (folded into cristae) Two compartments: (1) the intermembrane space (2) the matrix Location of mitochondrial complexes • Inner mitochondrial membrane: Electron transport chainATP synthase • Mitochondrial matrix:Pyruvate dehydrogenase complexCitric acid cycleFatty acid oxidation The outer membrane is permeable to small molecules and ions because it contains pore-forming protein (porin). The inner membrane is impermeable to ions and polar molecules. Contains transporters (translocases).
NADH FMN Fe-S Co-Q Fe-S cyt c1 cyt c cyt a cyt a3 O2 cyt b succinate FAD Fe-S THE ELECTRON TRANSPORT CHAIN Series of enzyme complexes (electron carriers) embedded in the inner mitochondrial membrane, which oxidize NADH2 and FADH2 and transport electrons to oxygen is calledrespiratory electron-transport chain (ETC). The sequence of electron carriers in ETC
High-Energy Electrons: Redox Potentials and Free-Energy Changes In oxidative phosphorylation, the electron transfer potentialof NADH or FADH2 is converted into the phosphoryl transfer potentialof ATP. Phosphoryl transfer potential is G°' (energy released during the hydrolysis of activated phos-phate compound). G°' for ATP = -7.3 kcal mol-1 Electron transfer potential is expressed as E'o, the (also called redox potential, reduction potential, or oxidation-reduction potential).
NADH FMN Fe-S Co-Q Fe-S cyt c1 cyt c cyt a cyt a3 O2 cyt b succinate FAD Fe-S E'o (reduction potential)is a measure of how easily a compound can be reduced (how easily it can accept electron). All compounds are compared to reduction potential of hydrogen wich is 0.0 V. The larger the value of E'o of a carrier in ETC the better it functions as an electron acceptor (oxidizing factor). Electrons flow through the ETC components spontaneouslyin the direction of increasing reduction potentials. E'o of NADH= -0.32 volts (strong reducing agent)E'o of O2 = +0.82 volts (strong oxidizing agent)
Important characteristic of ETC is the amount of energy released upon electron transfer from one carrier to another. This energy can be calculated using the formula: Go’=-nFE’o n – number of electrons transferred from one carrier to another; F – the Faraday constant (23.06 kcal/volt mol); E’o– the difference in reduction potential between two carriers. When two electrons pass from NADH to O2 : Go’=-2*96,5*(+0,82-(-0,32)) = -52.6 kcal/mol