Evolution and Complexity of GPCR Signaling Networks
690 likes | 843 Vues
This study reviews the evolution of G-protein coupled receptors (GPCRs) and their signaling networks. It explores how receptors interact with G-proteins and effectors, leading to a complex regulatory landscape that impacts various cellular functions. The text examines functional regions within GPCRs, highlighting G-protein interacting domains and ligand binding properties. By analyzing receptor chimeras, the study investigates differences in signaling pathways among similar receptors. It aims to elucidate the cross-talk within GPCR-mediated signaling systems through comprehensive molecular characterization.

Evolution and Complexity of GPCR Signaling Networks
E N D
Presentation Transcript
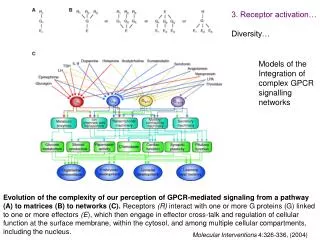
3. Receptor activation… Diversity… Models of the Integration of complex GPCR signalling networks Evolution of the complexity of our perception of GPCR-mediated signaling from a pathway (A) to matrices (B) to networks (C). Receptors (R) interact with one or more G proteins (G) linked to one or more effectors (E), which then engage in effector cross-talk and regulation of cellular function at the surface membrane, within the cytosol, and among multiple cellular compartments, including the nucleus. Molecular Interventions 4:326-336, (2004)
4. Functional regions within GPCRs: 4.1 G-protein interacting domains 4.2 ligand binding domain The three subfamilies of GPCRs are depicted with examples of their endogenous agonists. The binding modes of the orthosteric ligands for each receptor type are depicted by a green rectangle. The GPCR signals either by coupling to heterotrimeric G-proteins consisting of and subunits (which trigger a wide range of metabolic cascades and ion channel activities) or by direct association with effector molecules. AC, adenylyl cyclase; ATP, adenosine triphosphate; cAMP, cyclic adenosine monophosphate; PLC, phospholipase C; IP3, inositol-3,4,5-tris-phosphate; DAG, diacylglycerol.
4.Functional regions within GPCRs 4.1 G-protein coupling domain Using GPCR chimeras to identify Functional domains within receptors Two different dopamine receptors bind same agonist but they couple to different signalling cascade Mol Pharmacol. 2004 65:1323-32.
4. Functional regions within GPCRs Pharmacological characterization of GPCRs (A) Saturation isotherms: Determine the number of high affinity binding sites on the cell surface (B) Displacement analysis: Determine the affinity of the GPCR for different agonists and antagonists Binding properties of tritiated d[Cha4]AVP. Membranes from COS cells, expressing human V1b receptors (1-2 µg protein/assay), were incubated for 1 h at 37 °C with increasing amounts of tritiated d[Cha4]AVP (a) or with 1-2 nm of the radioligand with, or without (control), increasing amounts of unlabelled selective vasopressin antagonists (SR149415, SR49059, SR121463) (b). Journal of Neuroendocrinology16, 356-361
4.1 G-protein coupling domain… Analysis of vasopressin receptor chimeras Structure, ligand binding properties, and functional profile of wild type and mutant V1 /V2 vasopressin receptors. [ H]AVP saturation binding studies were carried out. Km and Bmax values are given as means ± S.E. of three independent experiments (PI, stimulation of PI hydrolysis; AC, stimulation of adenylyl cyclase). The symbols are defined as the percentage of maximum PI and cAMP responses induced by the wild type V1a and V2 receptor, respectively: ++++, 90-100%; +++, 80-90%; +, 10-30%; -, no significant response. J. Biol. Chem. 1996;271:8772
4.1 G-protein coupling domain… Analysis of the potency of Vasopressin chimeras to stimulate cAMP accumulation (Gs response) AVP-induced cAMP accumulation mediated by wild type V1a, V2 and hybrid V1a/V2 vasopressin receptors. Transfected COS-7 cells transiently expressing the different receptors were incubated in 6-well plates for 1 h at 37 °C with the indicated AVP concentrations, and the resulting increases in intracellular cAMP levels were determined. The data are presented as fold increase in cAMP above basal levels in the absence of AVP. Each curve is representative of three independent experiments, each carried out in duplicate. J. Biol. Chem. 1996;271:8772
4.1 G-protein coupling domain… Analysis of the potency of Vasopressin chimeras to stimulate Inositol-Phosphate (PI or lipid hydrolysis) accumulation (Gq response) Figure 3: AVP-induced stimulation of PI hydrolysis mediated by wild type V1a, V2 and hybrid V1a/V2 vasopressin receptors. Transfected COS-7 cells transiently expressing the various receptors were incubated in 6-well plates for 1 h at 37 °C with the indicated AVP concentrations, and the resulting increases in intracellular IP levels were determined. The data are presented as fold increase in IP above basal levels in the absence of AVP. Each curve is representative of three independent experiments, each carried out in duplicate. J. Biol. Chem. 1996;271:8772
4.1 G-protein coupling domain… Fine structure mapping of G-protein interacting region using deletion analysis Localization of FSHR Mutations and Functional Characterization of Mutant Receptors To characterize mutant FSHRs (A), COS-7 cells were transfected with the different constructs, and cAMP accumulation assays were performed as outlined in Materials and Methods. (B) Data obtained from three independent experiments, each performed in triplicate, are presented as -fold of basal cAMP levels of the FSHR(wt) (means ± SEM). Molecular Endocrinology 1999, 13: 181-190
4.1 G-protein coupling domain… Fine structure mapping of G-protein interacting region using alanine mutagenesis Functional Analysis of Conserved Basic Amino Acids within the LHR i2 Loop by Alanine Scanning Mutagenesis. To identify a cationic contact site for D564, all conserved basic amino acids within the i2 loop were replaced by A using a site-directed mutagenesis approach (A). The various LHR mutants were expressed in COS-7 cells, and cAMP accumulation assays were performed (B). Basal (open bars) and agonist-induced cAMP levels are presented as means ± SEM of two independent experiments, each carried out in triplicate. Molecular Endocrinology 1999, 13: 181-190
4. Functional regions within GPCRs 4.2 Ligand binding domain 3-D models Schematic models of ligand-receptor complexes for structurally diverse ligands interacting with GPCRs.
4. Functional regions within GPCRs 4.2 Ligand binding domain Using GPCR chimeras to identify Functional domains within receptors Two different receptors one binds FSH and the other binds LH Mol Pharmacol. 2004 Jun;65(6):1323-32.
4.2 Ligand binding domain… Structure of Cholecystokinin R eceptor Binding Sites and Mechanism of Activation/Inactivation by Agonists/Antagonists A wide repertoire of physiological effects of CCK and/or gastrin which mediated CCK1 and/or CCK2 receptors has been identified. Among actions of CCK which are mediated by the CCK1R, control of satiety, gallbladder contraction, pancreatic exocrine secretion, gastric pepsinogen and leptin secretions, gastric emptying and gut motility are the best known. Actions of CCK which occur through the CCK2R include modulation of anxiety and pain perception; these actions involve CCK2 receptors of the central nervous system. The wide spectrum of biological functions regulated by the CCK1R and CCK2R makes them candidate targets for a therapeutic approach in a number of diseases. This led a number of academic and pharmaceutical research groups to design specific and highly potent agonists and antagonists for those receptors. As for other G-protein-coupled receptors, the cloning of CCK receptor cDNAs and genes have stimulated generation of new biological models (transgenic animals, genetically modified cells ) and opened new avenues for research in the physiology, pathophysiology, molecular pharmacology and structure-function relationships of the receptors. Among these themes, delineation of CCK receptor binding sites represents a prerequisite for the understanding of the molecular basis for ligand recognition, partial agonism, ligand-induced traffiking of receptor signalling etc. Pharmacology & Toxicology 2002 91;313
4.2 Ligand binding domain… Criteria used to identify residues of CCKR binding site in an example of analysis of a putative ionic interaction between a positively charged aminoacid Y+ of the receptor binding site, and its negatively charged partner residues X in the ligand. Pharmacology & Toxicology 91:313, 2002
Schematic representations of the CCK1R with residues of the CCK binding site. Pharmacology & Toxicology 91:313, 2002
4.2 Ligand binding domain… Detailed understanding of residues involved in ligand binding allows detailed ligand/ Receptor modeling and permits the application of rational drug design to develop new agonists and antagonist that can be used as receptor specific drugs. Localizations of residues in biogenic amine receptors that are important for ligand binding (gray color) within a membrane topology model of the rat 5HT2A receptor (N terminus and most of C terminus are not shown). Pharmacol Ther. 2004 Jul;103(1):21-80
4. Functional regions within GPCRs Summary Generic diagram of sequence regions involved in post-translational modification and modulation of functions. Summary of findings based on receptor mutagenesis and construction of receptor chimeras for a variety of GPCR. Molecular Interventions 4:326-336, (2004)
5. Inactivation of GPCR signalling -Tachyphylaxis of the adrenomedulin (AM) receptor • Concentration-dependent increase in cAMP in response to AM (EC50 3.2±0.7 nM, filled squares) and CGRP at 1 µM (no effect, filled triangle) in Rat-2 fibroblasts. • Concentration-dependent attenuation of AM cAMP responses following preexposure to AM. Rat-2 cells were incubated with SFM or various concentrations (as indicated) of AM for 1 h followed by a washout period and a restimulation of 15 min with 10 nM AM Data are expressed as a percentage of the response in cells not preincubated with AM, that is, nondesensitised controls. Cells incubated with AM for 1 h were less able to elevate cAMP upon restimulation with AM when compared with cells incubated with SFM for the same time. • (c) The time course for AM receptor desensitisation in Rat-2 cells was determined by varying the preincubation time period (as indicated) of exposure to 100 nM AM. Cells were then washed and restimulated, and the cAMP response was measured. The cAMP response elicited by 10 nM AM was attenuated in a time-dependent manner. Cells preexposed to 100 nM AM for 1 min were capable of only 59.1±7% of the response of cells not preincubated with AM. After 2 h preincubation with 100 nM AM, the level of cAMP response was 19.3±2.2% of control stimulation. Regul Pept. 2003;112:139
5. Inactivation of GPCR signalling… GPCR Kinase and -arrestin-dependent desensitization and internalization of GPCRs Fergusson, 2001
5. Inactivation of GPCR signaling… Visulialization of GPCR internalization following receptor stimulation Translocation of ß-arrestin 2-GFP to the ß2-adrenergic receptor (ß2AR). HEK 293 cells stably overexpressing the ß2AR were transiently transfected with ß-arrestin 2-GFP. The distribution of ß-arrestin 2-GFP fluorescence was visualized by confocal microscopy before (−Iso) and after a 5 min treatment with isoproterenol (+Iso; is a ßAR agonist, 10−8, 10−6 M) at 37 °C. Before agonist-stimulation, ß-arrestin 2-GFP is uniformly distributed throughout the cytosol. Upon agonist addition, ß-arrestin 2-GFP translocates from the cytosol to the plasma membrane where it is found colocalizing with the receptor in punctuated areas of the plasma membrane. Prog Neurobiol. 2002 Feb;66(2):61-79
5. Inactivation of GPCR signaling… Model depicting the regulation of β2AR internalization, trafficking, dephosphorylation, and recycling Fergusson, 2002
5. Inactivation of GPCR signaling… Model depicting the regulation of GPCR internalization, trafficking, dephosphorylation, and recycling. Fig. 8. Pathways involved in desensitization and resensitization of GPCR signaling. Typically activation of a GPCR leads to (1) activation and inhibition of specific signaling pathways in the cell, (2) short-term desensitization mediated by phosphorylation of GPCRs by GRKs followed by β-arrestin binding to GPCRs that uncouple the receptor at the plasma membrane from the G-protein, (3) endocytosis of the receptor, followed by postendocytic sorting of the receptor either (4) back to the plasma membrane or (5) to lysosomes for degradation. Molecular Interventions 4:326-336, (2004)
5. Inactivation of GPCR signaling… Structure of GRKs Schematic representation of the domain architecture for GRK1-GRK7. The amino-terminal GPCR-binding domain of GRK1-GRK7 contains a conserved RGS domain. The plasma membrane targeting of each of the GRKs is mediated by distinct mechanisms that involves their carboxyl-terminal domains. GRK1 and GRK7 are farnesylated at CAAX motifs in their carboxyl termini. The carboxyl-terminal domains of GRK2 and GRK3 contain a βγ-subunit binding domain that exhibits sequence homology to a pleckstrin homology domain. The GRK5 carboxyl-terminal domain contains a stretch of 46 basic amino acids that mediate plasma membrane phospholipid interactions Pharmacol Rev. 2001;53:1-24.
5. Inactivation of GPCR signaling… Nature Reviews Molecular Cell Biology3; 639-650 (2002)
5. Inactivation of GPCR signaling… Regulators of G-protein Signalling (RGS) negatively regulate GPCR signalling N Extracellular Environment C GDP GTP + RGS Pi Downstream effectors
5. Inactivation of GPCR signaling… Prototypical RGS members Wieland and Mittmann 2003
5. Inactivation of GPCR signaling… Up-regulation of RGS4 desensitises endothelin-1 signalling in failing human myocardium In Congestive Hearts or Sepsis Also see increases in RGS1, 3, 16 Altered RGS expression leads to changes in GPCR signalling. Wieland and Mittmann 2003
6. GPCR dimerization: The Split Receptor Schematic representation of the wild-type human M2 and rat M3, the fragments M2-trunc and M3-tail, and the mutants M3-short and M2 (Asn404 to Ser) muscarinic receptors. The truncated fragment, M2-trunc, contains the amino-terminal domain, the first five hydrophobic transmembrane regions and the initial portion (56 amino acids) of the i3 loop of the wild-type muscarinic M2 receptor. The M3-tail fragment contains the final portion of the i3 loop (105 amino acids), the last two hydrophobic transmembrane regions, and the carboxy-terminal segment of the wild-type M3 muscarinic receptor. The short construct (M3-short) represents a receptor in which 196 amino acids of the i3 loop have been deleted; the remaining i3 loop is 43 amino acids long. The point mutant M2 (Asn404 to Ser) has the asparagine 404 replaced with serine.
6. GPCR dimerization… Figure 1 | Role of homo- and heterodimerization in the transport of G-protein-coupled receptors. When expressed alone, the GABABR1 (GBR1) receptor is retained as an immature protein in the endoplasmic reticulum (ER) of cells and never reaches the cell surface. By contrast, the GBR2 isoform is transported normally to the plasma membrane but is unable to bind GABA and thus to signal. When coexpressed, the two receptors are properly processed and transported to the cell surface as a stable dimer, where they act as a functional metabotropic GABAB receptor. Nat Rev Neurosci. 2001 Apr;2(4):274-86
6. GPCR dimerization… Molecular determinants of G-protein-coupled-receptor dimerization. Distinct intermolecular interactions were found to be involved for various G-protein-coupled receptors. Covalent disulphide bonds were found to be important for the dimerization of the calcium-sensing and metabotropic glutamate receptors. A coiled-coil interaction involving the carboxyl tail of the GBR1 and GBR2 receptors is involved in the formation of their heterodimer. Finally, for monoamine receptors such as the β2-adrenergic and dopamine receptors, interactions between transmembrane helices were proposed to be involved.
6. GPCR dimerization… Alternative three-dimensional models showing dimers of G-protein-coupled receptors. Two models have been proposed for the general three-dimensional organization of G-protein-coupled-receptor dimers. a | First is the domain-swapping model in which each functional unit within the dimer is composed of the first five transmembrane domains of one polypeptide chain and the last two of the other. Such a model is useful to rationalize the functional complementation observed when mutant or chimeric receptors are coexpressed. b | Second is the contact model in which each polypeptide forms a receptor unit that touches the other through interactions involving transmembrane domains five and six.
6. GPCR dimerization… Heterodimerization of CRLR and RAMP. HOMOTROPIC interactions between G-protein-coupled receptors (GPCRs) are not the only type of protein–protein interaction shown to influence their functional expression. Recently, a new class of membrane proteins that can interact with GPCRs and affect their activity profile has been identified. These new proteins were discovered while studying the expression of a complementary DNA that encoded a putative GPCR, which did not lead to the expression of a functional receptor. Specifically, a cDNA named calcitonin-receptor-like receptor (CRLR), which showed 55% overall identity to the calcitonin-receptor gene, was proposed to encode the receptor for the calcitonin-gene-related peptide (CGRP). However, various attempts to show that it was indeed the CGRP receptor failed because it was impossible to demonstrate any type of functional expression.
6. GPCR dimerization… Potential roles of G-protein-coupled receptor (GPCR) dimerization during the GPCR life cycle.(1) In some cases, dimerization has been shown to have a primary role in receptor maturation and allows the correct transport of GPCRs from the endoplasmic reticulum (ER) to the cell surface. (2) Once at the plasma membrane, dimers might become the target for dynamic regulation by ligand binding. (3) It has been proposed that GPCR heterodimerization leads to both positive (+) and negative (-) ligand binding cooperativity, as well as (4) potentiating (+)/attenuating (-) signalling or changing G-protein selectivity. (5) Heterodimerization can promote the co-internalization of two receptors after the stimulation of only one protomer. Alternatively, the presence of a protomer that is resistant to agonist-promoted endocytosis, within a heterodimer, can inhibit the internalization of the complex. G, G protein; L, ligand. EMBO Rep. 2004; 5:30-4.
6. GPCR dimerization… Summary of studies on hetero-dimerization in different GPCR classes and the possible functional roles of GPCR dimerization
6. GPCR dimerization… Summary of studies on hetero-dimerization in different GPCR classes and the possible functional roles of GPCR dimerization (cont’d)
Summary of factors effecting GPCR signalling The coupling of GPCRs with multiple G-proteins is selectively regulated at different levels. First, the nature of the response is dependent on the agonist used, which may selectively favour the coupling with a subset of G-proteins. In addition, when multiple couplings occur, several studies have demonstrated that the agonist elicited the responses with different potencies. Therefore, the involvement of multiple G-proteins is influenced by the concentration of agonist. Secondly, alterations in the expression or in the structure of the receptor have been shown to affect the coupling profile with multiple G-proteins. Thus, distinct coupling properties have been observed for related splice variants of a same receptor. Post-translational modifications, such as palmitoylation and phosphorylation, are also involved in the dynamic regulation of the G-protein coupling specificity. Many recent studies have shed light on the critical role played by GPCR-interacting proteins in determining the efficiency of coupling with distinct G-proteins. Finally, the availability of distinct G-proteins and the selective modulation of their expression, localisation, and activity contribute to determining the specificity of the intracellular signalling triggered after the receptor activation. Pharmacol Ther. 2003 99:25-44
Figure 1. HIV-1 fusion according to current models Env is composed of a surface subunit gp120 and transmembrane subunit gp41, which are non-covalently associated and then assembled as a trimer. The CD4 binding region of gp120 is exposed, but variable regions of gp120 screen important conserved structures involved in chemokine receptor interactions. Each gp41 molecule contains two alpha-helices that form a hairpin configuration, and the N-terminal hydrophobic fusion peptide is buried within the complex. gp120 binds to CD4 (A) and undergoes conformational changes that create or unmask the co-receptor binding site (B) and enable binding to the chemokine receptor (C). Structural changes are then induced in gp41 that extend the helical domains to form a ‘pre-hairpin intermediate’ (D). The hydrophobic fusion peptide inserts into the target cell membrane, causing gp41 to span between the virus and cell membranes. The gp41 helices then fold into a six-helix bundle, bringing together the N-terminal and C-terminal domains and thus the viral and cellular membranes (E). Contact between the membranes allows mixing of the outer leaflets followed by the development of a fusion pore (G). gp120 is omitted from panels F and G for the sake of clarity. Modified after Starr-Spires and Collman [6]. From: Shaheen: Curr Opin Infect Dis, Volume 17(1).February 2004.7-16 7. Alternative functions of GPCRs
The Nobel Prize in Physiology or Medicine 2004 B1. Odorant receptors "for their discoveries of odorant receptors and the organization of the olfactory system"
B1. Odorant receptors… The sense of smell: genomics of vertebrate odorant receptors.Olfactory receptor (OR) proteins interact with odorant molecules in the nose, initiating a neuronal response that triggers the perception of a smell. The OR family is one of the largest known mammalian gene families, with around 900 genes in human and 1500 in mouse. After discounting pseudogenes, the functional repertoire in mouse is more than three times larger than that of human. OR genes encode G-protein-coupled receptors containing seven transmembrane domains. ORs are arranged in clusters of up to 100 genes dispersed in 40-100 genomic locations. Each neuron in the olfactory epithelium expresses only one allele of one OR gene. The mechanism of gene choice is still unknown, but must involve locus, gene, and allele selection. The gene family has expanded mainly by tandem duplications, many of which have occurred since the divergence of the rodent and primate lineages. Interchromosomal segmental duplications including OR genes have also occurred, but more commonly in the human than the mouse family. As a result, many human OR genes have several possible mouse orthologs, and vice versa. Sequence and copy number polymorphisms in OR genes have been described, which may account for interindividual differences in odorant detection thresholds. Hum Mol Genet. 2002,11:1153
B1. Odorant receptors… Figure 32.2. The Main Nasal Epithelium. This region of the nose, which lies at the top of the nasal cavity, contains approximately 1 million sensory neurons. Nerve impulses generated by odorant molecules binding to receptors on the cilia travel from the sensory neurons to the olfactory bulb. Biochemistry.Berg, Jeremy M.; Tymoczko, John L.; and Stryer, Lubert
The olfactory system The olfactory epithelium contains millions of olfactory neurons, which send messages directly to the olfactory bulb of the brain. The olfactory receptor cells are the only neurons in the nervous system exposed directly to the external environment.
B1. Odorant receptors… (A) The combinatorial code of olfaction. Neurons expressing a given receptor can respond to more than one type of odorant (e.g. green receptor). Each odorant can elicit responses from several receptors, perhaps with different response amplitudes (here, the red receptor reacts strongly and the green receptor less strongly). Thousands of neurons expressing a given olfactory receptor are spread throughout one zone of the olfactory epithelium, but their axons converge on one or two glomeruli in the olfactory bulb. (B) Sources of phenotypic variation in olfaction. Individuals with different genotypes may (1) be homozygous for a given olfactory receptor, (2) express sequence variants with slightly different odorant-binding capabilities, (3) possess non-functional variants (hatched receptor) and/or (4) have duplicate gene copies, perhaps changing relative numbers of responsive neurons in the olfactory epithelium. Human Molecular Genetics, 2002, 11, 1153
B1. Odorant receptors… Genomic organization of olfactory receptor genes. Top: OR genes have a single main coding exon (black) and typically have several 5' untranslated exons. Alternate splicing is seen in many genes. Middle: OR genes are clustered in the genome in groups of 1 to over 100 genes (green arrows) and pseudogenes (red arrows) in both transcriptional orientations. Bottom: OR clusters are dispersed around the genome in more than 40 (mouse) or over 100 (human) locations.
Models of odorant receptor (OR) transcriptional regulation. (a) The short promoter model. The sequences immediately upstream of OR genes contain transcription factor binding sites sufficient to regulate receptor transcription. Which OR gene is expressed depends on the transcription factor(s) expressed in the cell. (b) The locus control region (LCR) model. Proximal promoters contain transcription factor binding sites necessary to drive specific receptor transcription, but are unavailable until factors binding to a distal LCR makes the region transcriptionally accessible. (c) The recombination model. OR genes are translocated by recombination or copied by gene conversion into a single active locus for expression. Transcription promoting factors are depicted as ovals. Trends Genet. 2002 Jan;18(1):29-34
B1. Odorant receptors… Figure 1. Gene-translocation mechanisms that activate one member of the multigene family. (a) DNA recombination. As seen in the mouse immunoglobulin (Ig) κ light-chain genes, DNA deletion brings a promoter carried by each variable (V) gene segment and the enhancer region between the joining (J) and constant (C) gene segments into proximity, thus activating the translocated gene. (b) Gene conversion. This is another gene-translocation mechanism that activates one particular member of the multigene family. A copy of the gene to be activated is transferred into the expression cassette located remotely from the gene cluster. This activation mechanism can be found in the yeast mating-type choice and antigenic variation in African trypanosomes. Abbreviations: P, promoter; E, enhancer. Trends Genet. 2004 Dec;20(12):648-53
Figure 32.5. The Olfactory Signal-Transduction Cascade. The binding of odorant to the olfactory receptor activates a signaling pathway similar to those initiated in response to the binding of some hormones to their receptors The final result is the opening of cAMP-gated ion channels and the initiation of an action potential.
Table 1 (Examples of Molecules in Different Odor Classes): B1. Odorant receptors…