Download

1 / 83
970 likes | 1.16k Vues
Explore the genetic basis of cancer, from mutations in cells to the development of tumors. Learn about the properties of cancer cells, how tumors grow, and the classification of cancers based on tissue types. Discover how genetic diseases differ from cancer and the importance of inherited cancer syndromes. Gain insights into the complexity of cancer development and why a single mutation is not enough to cause cancer.

E N D
Cancer; definition • Unlike free-living cells such as bacteria, which compete to survive, the cells of a multicellular organism are committed to collaboration. • To coordinate their behavior, the cells send, receive, and interpret an elaborate set of signals that serve as social controls, telling each of them how to act. As a result, each cell behaves in socially responsible manner, resting, dividing, differentiating, or dying as needed for the good of the organism. • In a human body with more than 1014 cells, billions of cells experience mutations every day, potentially disrupting the social controls. Most dangerously, a mutation may give one cell a selective advantage, allowing it to divide more vigorously than its neighbors and to become a founder of a growing mutant clone. • A mutation that gives rise to such selfish behavior by individual members of the cooperative can jeopardize the future of the whole enterprise. Repeated rounds of mutation, competition, and natural selection operating within the population of somatic cells cause matters to go from bad to worse. These are the basic ingredients of cancer: it is a disease in which individual mutant clones of cells begin by prospering at the expense of their neighbors, but in the end destroy the whole cellular society.
Cancer cells are defined by two heritable properties. they and their progeny • reproduce in defiance of the normal restraints on cell division • invade and colonize territories normally reserved for other cells. • An isolated abnormal cell that does not proliferate more than its normal neighbors does no significant damage, no matter what other disagreeable properties it may have; but if its proliferation is out of control, it will give rise to a tumor, or neoplasm—a relentlessly growing mass of abnormal cells
Benign As long as the neoplastic cells remain clustered together in a single mass, however, the tumor is said to be benign. At this stage, a complete cure can usually be achieved by removing the mass surgically. • Malignant A tumor is considered a cancer only if it is malignant, that is, only if its cells have acquired the ability to invade surrounding tissue. Invasiveness usually implies an ability to break loose, enter the bloodstream or lymphatic vessels, and form secondary tumors, called metastases, at other sites in the body. The more widely a cancer spreads, the harder it becomes to eradicate.
Cancers are classified according to the tissue and cell type from which they arise • Cancer arising from epithelial cells are termed carcinomas • Those arising from connective tissue or muscle cells are termed sarcomas. • Cancers that do not fit in either of these two broad categories include the various leukemias, derived from hemopoietic cells, and cancers derived from cells of the nervous system.
Cancer Is a Unique Type of Genetic Disease • The classical genetic diseases are typically monogenic in nature; they are caused by a single faulty gene. In some cases, the inheritance of a defective gene is both necessary and sufficient to cause disease, and the incidence is exactly as predicted by the Mendelian laws of inheritance. (e.g Sickle cell anemia) cancer can be inherited as a monogenic trait. Large, extended families have been identified in which individuals in multiple generations develop etiologically related types of cancer at a high rate. Such families have been used to define cancer syndromes and to isolate the genes that underlie cancer susceptibility. While inherited cancer syndromes have provided a wealth of information as to the relationships between specific genes and cancer, they are relatively rare.
Thus, a cardinal feature of aneuploidy could be experimentally separated from the underlying process that causes CIN. • Aneuploidy is a reflection of an ongoing cellular process • Aneuploidy does not in and of itself cause instability, but rather may result from instability • Aneuploidy, in the forms examined in this analysis of prototypical colorectal cancer cells, is a dominant phenotype.
A Single Mutation Is Not Enough to Cause Cancer • An estimated 1016 cell divisions take place in a normal human body in the course of a lifetime; in a mouse, with its smaller number of cells and its shorter life span, the number is about 1012. • Even in an environment that is free of mutagens, mutations will occur spontaneously at an estimated rate of about 10-6 mutations per gene per cell division—a value set by fundamental limitations on the accuracy of DNA replication and repair. • Thus, in a lifetime, every single gene is likely to have undergone mutation on about 1010 separate occasions in any individual human being. Among the resulting mutant cells one might expect that there would be many that have disturbances in genes that regulate cell division and that consequently disobey the normal restrictions on cell proliferation. From this point of view, the problem of cancer seems to be not why it occurs but why it occurs so infrequent.
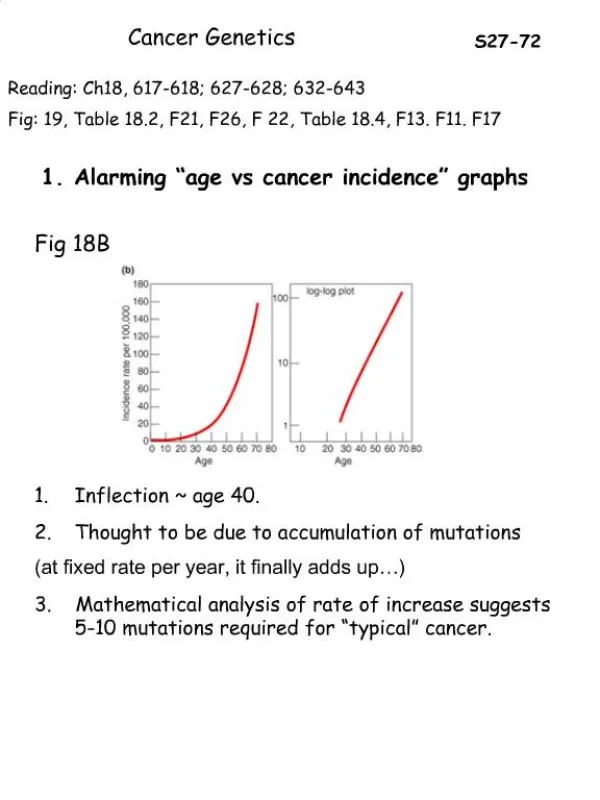
Clearly, if a single mutation were enough to convert a typical healthy cell into a cancer cell that proliferates without restraint, we would not be viable organisms. • Many types of evidence indicate that the genesis of a cancer typically requires that several independent, rare accidents occur in the lineage of one cell. • One such indication comes from epidemiological studies of the incidence of cancer as a function of age. If a single mutation were responsible, occurring with a fixed probability per year, the chance of developing cancer in any given year should be independent of age. • In fact, for most types of cancer the incidence rises steeply with age—as would be expected if cancer is caused by a slow accumulation of numerous random mutations in a single line of cells.
A tumor develops through repeated rounds of mutation and proliferation, giving rise eventually to a clone of fully malignant cancer cells. At each step, a single cell undergoes a mutation that enhances cell proliferation, so that its progeny become the dominant clone in the tumor. Proliferation of this clone then hastens occurrence of the next step of tumor progression by increasing the size of the cell population at risk of undergoing an additional mutation. • Initiation • Progression • Metastasis
Why are so many mutations needed? • One reason is that cellular processes are controlled in complex and interconnected ways; cells employ redundant regulatory mechanisms to help them maintain tight and precise control over their behavior. Thus, many different regulatory systems have to be disrupted before a cell can throw off its normal restraints and behave defiantly as a malignant cancer cell. • In addition, tumor cells may meet new barriers to further expansion at each stage of the evolutionary process. • For example, oxygen and nutrients do not become limiting until a tumor is one or two millimeters in diameter, at which point the cells in the tumor interior may not have adequate access to such necessary resources. Each new barrier, whether physical or physiological, must be overcome by the acquisition of additional mutations
Six Key Properties Make Cells Capable of Cancerous Growth • They disregard the external and internal signals that regulate cell proliferation. • They tend to avoid suicide by apoptosis. • They circumvent programmed limitations to proliferation, escaping replicative senescence and avoiding differentiation. • They are genetically unstable. • They escape from their home tissues (that is, they are invasive). • They survive and proliferate in foreign sites (that is, they metastasize).
Cancer genes are distinct alleles of normal genes that arise as a result of mutation. • Germ cells mutation (0.1%-10%) • Somatic cells mutation (most frequent) • Viral infection
Mutations Alter the Human Genome • Microdeletion • Microinsertion • Gross deletion • Gross insertion • Inversion • Translocation • Repeated sequence
Genes and Mutations • Gene and locus: >1000-100000 • Nucleotides : A, T, C, G • Open reading frame • Mutation in splice sites : 10% • Mutation in intron • Mutation in Poly A site • Mutation of stop codon • New definition for gene 20%
The Majority of Cancer Cells Are Aneuploid • One of the most readily observable traits of cancer cells is an abnormal number of chromosomes. • While normal human somatic cells invariably contain 23 pairs of chromosomes, the cells that compose tumors often deviate significantly from this diploid complement. A cell that has a number of chromosomes that is not a multiple of the haploid number is defined as aneuploid. • A chromosome complement of between 60 and 90 chromosomes is frequently observed. • This number varies from cell to cell even within a single tumor.
… • Aneuploidy can be visually linked to mitosis, the phase of the cell cycle during which chromosomes condense, are aligned with respect to the division plane and are then pulled to opposing poles of the mitotic spindle. • The features of aneuploidy in cancer cells were first described by the German pathologist David Paul von Hansemann a decade after the discovery of chromosomes in the late 1870s. Note: While abnormal mitoses and chromosome complements had been observed in cancer tissues before, the prevailing thinking had held that these features were the result of fusions between neighboring tumor cells.
The hypothesis put forth by von Hansemann was extended and popularized several years later by the German biologist Theodor Boveri, who was among the first to link chromosomes with the functional units of heredity. Boveri noted that mitotic spindles in cancer cells often appeared to be multipolar, and suggested that an underlying mechanical defect that led to abnormal chromosome numbers was a pathological feature of the cancer cell.
In addition to these numerical abnormalities, the chromosomes in aneuploid cells commonly have structural aberrations that are rarely observed in normal cells. • The structural abnormalities associated with aneuploidy include cytogenetically apparent: • Translocations • Deletions • Inversions • Duplications.
… • Modern cytogenetic techniques vividly reveal both the numerical and structural abnormalities that define aneuploidy
What is the meaning of the striking chromosomal abnormalities that are found so frequently in cancer cells? • Is aneuploidy causally involved in cancer? • Is a consequence of hyperactive cell division?
The descriptions of aneuploidy provided by von Hansemann and Boveri suggested that aneuploidy might be a manifestation of an underlying defect in mitosis. • An alternative interpretation is that aneuploidy arises by some other means, and that mitosis is simply more likely to fail in the presence of too many chromosomes. • Christoph Lengauer (1990) • fluorescence in situ hybridization(FISH) • measured the rates at which chromosomes are lost and gained in colorectal cancer cells during long term culture
Cancer cells that were aneuploid tended to gain and lose individual chromosomes at the relatively high rate of 0.01 per chromosome per cell division . The increased rate of chromosome gains and losses in aneuploid cells was termed chromosomal instability, often abbreviated as CIN.
Animal models also confirm that a single one of these genetic alterations is insufficient to cause cancer: when genetically engineered in a mouse, a single such mutation typically produces mild abnormalities in tissue growth, followed occasionally by the formation of randomly scattered benign tumors; but the vast majority of cells in the mutant animal remain non-cancerous
Each cancer has characteristics that reflect its origin • One clear demonstration of clonal evolution comes from analysis of the chromosomes in tumor cells. • Chromosomal aberrations and rearrangements are present in the cells of most common cancers. • In almost all patients with chronic myelogenous leukemia (CML) , for example, leukemic white blood cells can be distinguished from normal cells by a specific chromosomal abnormality: the so-called Philadelphia chromosome, created by a translocation between the long arms of chromosomes 9 and 22.
Many other lines of evidence, from a variety of cancers, point to the same conclusion: most cancers originate from a single aberrant cell • As a result of a random process that occurs in the early embryo, practically every normal tissue in a woman's body is a mixture of cells with different X chromosomes heritably inactivated (indicated here by the mixture of red cells and gray cells in the normal tissue). • When the cells of a cancer are tested for their expression of an X-linked marker gene, however, they are usually all found to have the same X chromosome inactivated. This implies that they are all derived from a single cancerous founder cell.
Oncogenes • Oncogenes are the altered forms of normal genes-protooncogenes- that have key roles in cell growth and differentiation pathways. • In normal mammalian cells are sequences of DNA that are homologous to viral oncogenes; these that are named proto-oncogenes or cellular oncogenes. • Although the terms proto-oncogene and cellular oncogene are often used interchangeably, strictly speaking proto-oncogene is reserved for the normal gene and cellular oncogene, or c-onc, refers to a mutated proto-oncogene, which has oncogenic properties such as the viral oncogenes, or v-onc. • At least 50 oncogenes have been identified.
Types of Oncogene • Growth Factors • Growth Factor Receptors • Intracellular Signal Transduction Factors • DNA-Binding Nuclear Proteins • Cell-Cycle Factors
Relationship Between C-ONC and V-ONC • Cellular oncogenes are highly conserved in evolution, suggesting that they have important roles as regulators of cell growth, maintaining the ordered progression through the cell cycle, cell division, and differentiation. • Retroviral oncogenes are thought to acquire their dominant transforming activity during viral transduction through errors in the replication of the retrovirus genome following their random integration into the host DNA. The end result is a viral gene that is structurally similar to its cellular counterpart but is persistently different in its function.
Identification of Oncogenes • Identification of Oncogenes at Chromosomal Translocation Breakpoints • Chronic Myeloid Leukemia(CML): • Philadelphia, or Phi. Chromosome; The Phi is a tiny chromosome that is now known to be a chromosome 22 from which long arm material has been reciprocally translocated to and from the long arm of chromosome 9;t(9;22)(q34;qll). • This translocation has been found to transfer the cellular ABL (Abelson) oncogene from chromosome 9 into a region of chromosome 22 known as the breakpoint cluster, or BCR region, resulting in a chimeric transcript derived from both the c-ABL (70%) and the BCR genes. This results in a chimeric gene expressing a fusion protein consisting of the BCR protein at the amino end and ABL protein at the carboxy end, which is associated with transforming activity.
Burkitt Lymphoma • An unusual form of neoplasia seen in children in Africa is a lymphoma that involves the jaw, known as Burkitt lymphoma, named after Dennis Burkitt, a medical missionary who first described the condition in the late 1950s. • Chromosomal analysis has revealed the majority (90%) of affected children to have a translocation of the c-MYC oncogene from the long arm of chromosome 8 on to heavy (H) chain immunoglobulin locus on chromosome 14. • Less commonly the MYC oncogene is translocated to regions of chromosome 2 or 22, which encode genes for the kappa (K) and lambda (A.) light chains, respectively. • As a consequence of these translocations, MYC comes under the influence of the regulatory sequences of the respective immunoglobulin gene and is overexpressed 10-fold or more.
Oncogene Amplification • Proto-oncogenes can also be activated by the production of multiple copies of the gene or what is known as gene amplification, a mechanism known to have survival value when cells encounter environmental stress. • E.g: leukemic cells are exposed to the chemotherapeutic agent methotrexate, • In mammals the amplified sequence of DNA in tumor cells can be recognized by the presence of small extra chromosomes known as double-minute chromosomes or homogeneously staining regions of the chromosomes. These changes are seen in approximately 10% of tumors and are often present more commonly in the later rather than the early stages of the malignant process. • MYC, N-MYC, EBR-B2 and cyclin D1
What Is a Tumor Suppressor Gene? • A tumor suppressor gene is a type of cancer gene that is created by a loss-of- function mutation. • In contrast to the activating mutations that generate oncogenic alleles from proto- oncogene precursors, tumor suppressor genes and the proteins they encode are functionally in activated by mutations.
Tumor suppressor gene s typically control processes that are fundamental to the maintenance of stable tissue compartments.
… • Mutations that convert proto-oncogenes to oncogenes are: • Single nucleotide substitutions • Amplifications • Chromosomal rearrangements • Nonsense mutations • Frameshift mutations • Splice site mutations Common in Tumor suppressor Genes
The Discovery of Recessive Cancer Phenotypes • The first cancer genes to be discovered were oncogenes. • For a time it was accordingly believed that the cancer phenotype resulted primarily from activating mutations that led to gains of function. • In a 1969 study, Harris adopted a novel approach to study the genetic factors that were involved in cancer cell phenotypes.
Previously, it had been established that cells of different types could be experimentally fused, and thereby made to share their genes . • Among the phenotypes that many cultured cancer cells share is an ability to grow into tumors when implanted into mice, a property is known as tumorigenicity. • Tumorigenicity is an experimental trait that is believed to be reflective of the malignant nature of the tumor from which cultured cancer cell was derived.
The investigators concluded that tumorigenicity was a recessive trait that could be suppressed by the transfer of a specific chromosome, and perhaps even by the transfer of a single dominant gene.
Retinoblastoma and Knudson’s Two-Hit Hypothesis • Retinoblastoma is a tumor of the eye that arises from immature cells within the retina • Relatively rare tumor • Incidence : 1 in 20,000 live births • Sporadic tumors • Heritable tumors
Retinoblastoma occurs during infancy and early childhood, and accounts for 11 % of all cancers that occur in the first year of life. • Ninety percent of patients have no family history of the disease. • The cells of the retina are nearly fully differentiated at birth and have a limited capacity for further proliferation. • The early differentiation of the blast cells of the retina provides, in effect, a temporal window of cancer susceptibility. • Somatic mutations that occur within this proliferative window can potentially contribute to the subsequent growth of tumors
… • Retinoblastomas that occur in individuals with a family history of the disease frequently affect both eyes and exhibit multifocal tumors in a single eye. • However, the more common presentation, which accounts for about two thirds of all cases, is a tumor that is unifocal and restricted to one eye. • The bilateral form of the disease is, on average, diagnosed at 1 year of age. • The unilateral cases peak at about 2 years. Rare cases have been reported in older children and even in young adults.
Knudson’s two-hit hypothesis : The rate limiting event is the somatic mutation, which occurs at random and at a very low frequency during the limited temporal window of susceptibility
Chromosomal Localization of the Retinoblastoma Gene • Based on epidemiology alone, the two hits required for retinoblastoma could have been accounted for by mutations in two separate genes, perhaps two oncogenes that were sequentially activated. • However, the experiments by Harris had shed light on a completely novel and uncharacterized type of cancer gene that was recessive in nature. • Knudson suggested that both hits could occur in the two alleles of a single locus, and thereby cause the total inactivation of a recessive cancer gene. • Knudson’s hypothesis was supported and extended with the cloning and identification of the retinoblastoma susceptibility gene, RB1 .
Retinoblastoma tumor cells had no known biochemical or signaling defect. • There was therefore no basis to suspect that the causative gene might encode any particular enzyme or regulatory protein. • Ultimately, the retinoblastoma gene was not identified by virtue of its function, but rather on the basis of its location. • The first clue as to the location of the retinoblastoma locus arose from cytogenetic studies in the late 1970s.
Documents : • Microscopically visible deletions within one copy of chromosome 13 were observed in the normal blood cells of a small proportion of individuals with the inherited form of the disease. • While the extent of the constitutional deletion varied among retinoblastoma- prone individuals, one particular chromosomal band on the long arm of chromosome 13, designated 13q14, was consistently missing.