TRASCRIZIONE NEGLI EUCARIOTI
170 likes | 712 Vues
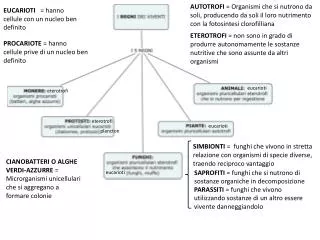
TRASCRIZIONE NEGLI EUCARIOTI. Due tappe nella regolazione dell'inizio della trascrizione. Attivazione trascrizionale per interazione diretta. Attivazione trascrizionale tramite complessi "mediatori". Il dominio "elica-giro-elica" (HTH). Il complesso d'inizio di Pol-II.

TRASCRIZIONE NEGLI EUCARIOTI
E N D
Presentation Transcript
Il termine splicing (in italiano letteralmente "saldatura") indica uno dei processi maturazione del trascritto primario dei geni discontinui. Il macchinario della sintesi proteica cellulare è in grado di tradurre solo gli RNA messaggeri che contengono segmenti continui di codoni, non ha modo di identificare ed evitare un blocco di sequenze non codificanti. Quindi, i trascritti primari dei geni devono avere i loro introni rimossi prima che avvenga la traduzione in proteine. È proprio attraverso lo splicing dell'RNA che gli introni vengono rimossi dal pre-mRNA. Questo processo converte il pre-mRNA in un messaggero maturo e deve essere molto preciso per evitare la perdita o l'aggiunta anche di un singolo nucleotide nei punti in cui gli esoni vengono uniti.
Chimicamente lo splicing avviene per due successive reazioni, grazie alle quali i legami vengono rotti e altri nuovi formati. Queste reazioni sono mediate da un meccanismo molecolare che coinvolge più componenti, chiamato spliceosoma. Le sue dimensioni sono simili a quelle di un ribosoma ed è composto da circa 150 proteine e da subunità. Sono proprio queste ultime a riconoscere, con molta probabilità, le sequenze ai confini introne-esone ed a partecipare alla catalisi della reazione. Le subunità sono chiamate ribonucleoproteine nucleari (snRNP, small nuclear ribonuclear protein). Le snRNP rivestono tre ruoli nello splicing: riconoscono il sito di splicing 5' e il punto di ramificazione; portano questi siti vicini quando occorre e catalizzano il taglio e la giunzione dell'RNA. Sono proprio queste interazioni e i successivi riarrangiamenti che ne conseguono che guidano la reazione di splicing e contribuiscono alla sua precisione.
Autosplicing Alcuni rari introni sono capaci di "autocatalisi", ovvero la parte intronica dell'RNA trascritto è un "ribozima" che catalizza la propria escissione. Questo meccanismo, definito "autocatalitico" perché può avvenire anche in completa assenza di proteine, riguarda introni di alcuni geni degli organelli, dei geni nucleari codificanti l'rRNA di qualche eucariote e di qualche gene procariotico.
Questa ricorrenza non è straordinaria. Gli introni possono essere escissi in maniere diverse, è il caso dello splicing alternativo - cioè il processo attraverso il quale, mediante un diverso arrangiamento degli esoni, da uno stesso gene possono derivare diverse proteine, dette isoforme. Circa l'60% dei geni degli Eucarioti superiori subisce lo splicing alternativo. Sull'mRNA è possibile individuare dei siti di splicing "forti" e altri siti "deboli"; questi ultimi sono dei siti criptici ossia siti che normalmente non sono riconosciuti come siti di splicing e che invece,vengono identificati come tali, in particolari condizioni. Il riconoscimento di tali siti permette la produzione di proteine differenti a partire dallo stesso mRNA.
Sono generalmente riconosciute cinque modalità di splicing alternativo. Salto dell'esone: un esone può essere eliminato dal trascritto primario. Questa è la modalità più comune di splicing nel pre-mRNA dei mammiferi Esone mutualmente esclusivo: solo uno di due esoni viene mantenuto nell'mRNA maturo, non entrambi Sito di taglio alternativo 5' : viene usato un sito di taglio al 5' alternativo, cambiando l'estremità 3' dell'esone a mont Sito di taglio alternativo 3' : viene usato un sito di taglio al 3' alternativo, cambiando l'estremità 5' dell'esone a valle Introne trattenuto: i siti di taglio di un introne possono non essere riconosciuti. In questo caso l'introne non viene eliminato dal trascritto di mRNA. Se l'introne trattenuto si trova nella regione codificante, esso non deve alterare la cornice di lettura degli esoni. Se avviene il cambiamento di quest'ultima, esso potrebbe generare una proteina tronca o non funzionale