
Nukleosomet
Nukleosomet. Forskjellige modeller for 30 nm-fibre. Ikke-Waston-Crick-basepar. Parring mellom adeninrester i krystallstrukturen av 9-metyladenin Hoogsteen-parring mellom adenin- og tyminrester i krystallstrukturen av 9-metyladenin/1-metyltymin Hypotetisk parring mellom cytosin- og tyminrester.
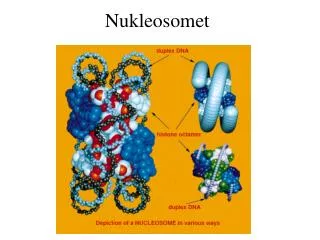

Nukleosomet
E N D
Presentation Transcript
Ikke-Waston-Crick-basepar • Parring mellom adeninrester i krystallstrukturen av 9-metyladenin • Hoogsteen-parring mellom adenin- og tyminrester i krystallstrukturen av 9-metyladenin/1-metyltymin • Hypotetisk parring mellom cytosin- og tyminrester
IR-spektrometri i upolare løsemidler viser hydrogenbindinger
Skjematisk oversikt over sekundærstrukturen til tRNA: kløverbladstrukturen.
The structure of the RNA of B. subtilis RNase P.. (a) Predicted secondary structure with specificity domain drawn in various colors and catalytic domain is black
The structure of the RNA of B. subtilis RNase P. (b) The X-ray structure of the specificity domain in which its various segments are colored as in Part a.
16S-rRNA-sekundærstruktur (E. coli ogarchae, pattedyr, mitokondrier)
Strukturen av et hammerhode-ribozym Venstre: Sekundærstruktur hvor prikkede linjer representerer Watson-Crick-basepar og ikke-WC-basepar representeres med heltrukne blå linjer Midt: RNA rød-blå, DNA gul-grønn Høyre: Skjematisk diagram av ribozymet
Forskjellige systemer for skjematisk fremstilling av RNA-strukturer
1958: the central dogma • DNA dirigerer replikasjon (DNA til DNA) og transkripsjon (DNA til RNA) og RNA dirigerer protein syntese (translasjon)
Disposisjon • Hvordan foregår transkripsjon? • Proteiner og andre komponenter • Prosesser og mekanismer • Medikamenter • Logikk - hva forventes - hva finner vi? • Hvordan reguleres transkripsjon • Logikk - hvilke angrepsmåter er sannsynlige? • Noen eksempler på ulike strategier
4 etapper • Binding • Initiering • Elongering • Terminering
Enzymet = RNA polymerase • Oppdaget 1960 • Samuel Weiss og Jerard Hurwitz • RNAn + NTP + (Mg++ + templat) = RNAn+1 + PPi • I bakterier - et enzym • Med unntak av primase • Noen fag koder for egne RNA polymeraser • Polypeptid sammensetning • 449 kDa • a2bb´s
Flere eukaryote RNA pol ansvarlig for ulike RNA • RNA polymerase I • rRNA - en promoter • RNA polymerase II • mRNA - komplekse promotere • RNA polymerase III • 5S RNA og tRNA - intrageniske promotere
RNA Polymerase Subunitsa. Page 1232
3D struktur • E.coli RNA pol T7 RNA polymerase
3D av Eukaryot RNA pol • Gjær RNA polymerase (lav resolusjon)
yRNAPII yRNAPII
An electron micrograph of E. coli RNA polymerase (RNAP) holoenzyme attached to various promoter sites on bacteriophage T7 DNA. Page 1222
X-Ray structure of Taq RNAP core enzyme.. Page 1224 a subunits are yellow and green, b subunit is cyan, b¢ subunit is pink, w subunit is gray
X-Ray structure of Taq RNAP. (b) The holoenzyme viewed as in Part a. Page 1224
Model of the closed (RPc) complex of Taq RNAP with promoter-containing DNA extending between positions –60 and +25. Page 1225
Model of the open (Rpo) complex of Taq RNAP with promoter-containing DNA showing the transcription bubble and the active site. Page 1225
Bindingens logikk - hva kan vi forvente? • Sekvensspesifikk binding • Sterkere binding til promoter enn til DNA generelt • Sterk binding gir effektiv RNA syntese, mens svak binding gir lavere nivåer av mRNA • Kjedeseparasjon før RNA-syntese
Promoterbinding • RNA pol danner sterke kompleks med promotere • Kd = 10-14M • DNase-proteksjon -20 til +20 • To konserverte områder oppstrøms • -10 region (Pribnow-box) TATAAT • -35 region (optimalt TTGACA) • Initieringsregion: +1 alltid A (CAT) eller G (CGT) • Sigma er bestemmende for spesifisitet
Initiering - hva er sekvens-signalet? • To konserverte regioner
RNA NTP 3´-OH Dannelse av åpent kompleks- separasjon av tråder • Området fra -9 til +2 åpnes • Evidens DMS-footprinting
Initieringens logikk - hva kan vi forvente? • Enzymatisk reaksjon • NTP + Mg++ + templat = RNA + PPi • Energi fra hvor? • Retning på kobling?
Uventet: abortiv initiering • Initiert RNA syntese ofte abortert etter 2-9 nukleotider • Oppfylling av RNA kanal - før overgang til elongeringskonformasjon
Medikament: Rifampicin inhiberer prokaryot transkripsjon • Streptomyces m. produkt = rifamycin B • Semisyntetisk rifampicin • Inhiberer spesifikt prokaryot transkripsjon via binding til b-subenhet og lager et bundet men blokkert enzym
Elongeringens logikk - hva kan vi forvente? • Kompleks av prosessiv type • som forblir værende i samme kompleks og forlenger det RNA som ble initiert • Topologisk nøtt
Retning av polymerisering Her vil innmerking holde seg lenge selv om hot GTP erstattes med kald Her vil innmerking raskt forsvinne når hot GTP erstattes med kald
Medikament: cordycepin (3-deoksyadenosin) • Blokkerer bakteriell RNA syntese • Virker ved inkorporering og kjedeterminering
Topologisk konsekvens av polymerisering • Hva er mest hensiktsmessig å snurre? RNA polymerasen - NEI DNA templaten - JA
Transkripsjon leder til supercoiling • Når uparet transkripsjonsboble beveger seg, dannes • Foran: positiv supercoiling (tettere tvunnet) • Plasmid blir positivt supercoilet i gyrasemutant • Bak: negativ supercoiling (undertvunnet) • Plasmid blir negativt supercoilet i topo I-mutant
Hvor rask skjer elongering? Hvor mye feil? • In vivo: 20-50 nt per sek • Initiering opp til 1x per sek • Fidelitet: feilinkorporering 1: 104
Medikament: det interkalerende middel Actinomycin D • Binder sterkt via interkalering og blokkerer både transkripsjon og replikasjon • Interkalering via phenoxazone ring • Minor groove binding av peptid del