Kromoszóma és replikáció
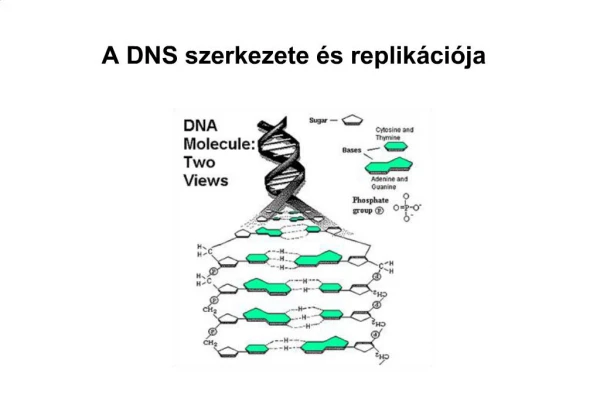
Kromoszóma és replikáció. A DNS szerkezete A baktérium kromoszóma szerveződése DNS replikáció Kromoszóma szegregáció. Bázisok. Bázispárok. A DNS szerkezete Váz 2 polinukleotid szál „jobbos” hélix, egymásba tekert, szét nem választható (plektoném) dupla hélix


Kromoszóma és replikáció
E N D
Presentation Transcript
Kromoszóma és replikáció • A DNS szerkezete • A baktérium kromoszóma szerveződése • DNS replikáció • Kromoszóma szegregáció
A DNS szerkezete Váz 2 polinukleotid szál „jobbos” hélix, egymásba tekert, szét nem választható (plektoném) dupla hélix A dupla hélixnek két árka, van: nagy és kicsi, átmérője 2 nm Mindegyik lánc d-2’ dezoxiribóz cukor alegységből áll, melyet foszfát csoportok kötnek össze 3’, 5’ foszfodiészter kötéssel Hosszú el nem ágazó polimer Az egyes bázisok a a cukor alegységekhez -N-glikozidos kötéssel kapcsolódnak A két szál antiparalel, az egyik szabad 5’ - 3’ irányú, míg a másik 3’ – 5’ irányú A bázisok a tengelyre merőlegesen helyezkednek el A spirál közötti teret töltik ki, A-T 2 hidrogénhíd, G-C 3 hidrogénhíd A bázisok távolsága (függőleges) 0,34 nm=3,4 Å 10 bp/fordulat, a B-formában (1 fordulat 3,4 nm Pirimidinnel szemben purin bázis
Különböző DNS szerkezetek „A”-DNS A bázisok planáris elhelyezkedés helyett ~ 30º-ban megdőlt Nagyobb feltekeredés 1 teljes fordulat a hélixben 11 bp A kis árok kinyílik Dupla szálú RNS-ben és RNS-DNS heteroduplexben főleg „A”-forma „Z”-DNS Ugyanaz a bázispárosodás A bázisok sztereokémiai konfigurációja syn és anti felváltva Syn a bázis síkja a cukormolekulából kifele mutat Anti a bázis síkja cukor síkja fölött helyezkedik el Következmény: dupla szálú balfordulatos DNS hélix Főleg pirimidin/purin monoton váltakozó szekvenciában alakul ki (A/T esetén nem) Mindent vagy semmi elv vagy B, vagy Z forma A DNS polimeráznak rossz szubsztrátja A B formával szemben megfelelő antigenitással rendelkezik, így mutatták ki Natív DNS molekulában is előfordul ?? (vagy az antitest alakítja ki ezt a szerkezetet) Lehet szabályozó szerepe E. coli 18 kb-onként 1 Z-DNS szakasz
Az előző szerkezet szerint milyen formák lehetnek (DNS toplógia) • Lineáris (nick egy helyen nincs kovalens kötés, megoldó funkció) • Relaxált cirkuláris • Szuperhelikális
A genom szerveződése Eukarióta Kromoszómába rendeződve Hiszton fehérjék H1, H2A, H2B, H3, H4 A H1 kivételével oktamer cilinder keletkezik, mely köré ~ 146 bp hosszú DNS molekula tekeredik Ez a nukleoszóma A nukleoszómák között 50-60 bp spacer van,amelyet H1 fehérje takar • A Saccharomyces (eukarióta) esetén a nukleoszómák közötti rész 20 bp • A H1 szerepe itt nem tisztázott • 16 kromoszómája van (200 kb-2,2 Mb)
A prokarióta genom Kezdetben azt gondolták, hogy a baktériumoknak nincs (az eukarióta kromoszómához hasonlítható) kromoszómájuk, ezért azt genofornak nevezték Van hasonlóság Autoradiográfiával kimutatják a coli cirkuláris genommal rendelkezik (mint a genetikai térkép) A coli DNS-e kb. 1 mm hosszú, kovalensen zárt kör, A sejt 1 m, azaz jól össze van pakolva 1000x-es különbség Intakt DNS-t izoláltak, autoradiográfiával nukleoid-ot mutatnak ki (lásd következő ábra) A nukleoid magja kettős és egyszálú DNS, RNS, fehérje A sűrűsége megfelel az emlős metafázisú kromoszómának A DNS hurkokba szerveződik A hurkok szuperspirális DNS-ből állnak A hurkok önállóak egy-egy nick csak egy rész szuperspiralitását oldja meg, domén szerkezet Kb. 50 hurokból, doménből áll A HU fehérjék (kisméretű bázikus) szerepe fontos lehet, kondenzálja a DNS-t, stimulálja a replikációt, H fehérjék szerepe nem ismert protaminhoz hasonlít az aminosav sorrendje
A sejtből izolált natív DNS szuperspirális • ~ 15 normál fordulatra jut egy szuperspirális fordulat • Nem magától alakul ki, topizomeráz enzimek alakítják ki • A szuperspiralitás lehet pozitív (jobbos) és negatív (balos) • Pozitív a mintha a DNS spirálját tovább feszítenénk • Negatív mintha a DNS spirálját ki szeretnénk tekerni • Az eddig izolált nukleoidok mind negatív szuperhelicitást mutatnak
DNS replikáció A DNS molekula in vivo szintézise Eukarióták és prokarióták között lényegében hasonlóan, az E. coli rendszere a legismertebb Watson/Crick modellből posztuláció, hogy a két szál szétválik és mindkét szál komplementer szálja megszintetizálódik Kísérletes bizonyíték Meselson és Stahl-tól a DNS molekula replikációja szemikonzervatív E. coli sejtek 15N izotópot tartalmazó táptalajon DNS izolálás Ultracentrifugálás a normál 14N forráson tartott coli DNS-nél nehezebb E. coli sejtek normál táptalajon DNS izolálás különböző szaporodási fázisban Ultracentrifugálás,a 14N-t és 15N izotópot tartalmazó sáv közötti DNS, amely egyenlő arányban tartalmazza a két N izotópot A keletkező DNS duplex egyik szála a régi, másik szála újonnan szintetizált A folyamatot katalizáló enzim a polimeráz (gyűjtőnév)
A DNS polimeráz az egyszálú DNS-ből kiindulva nem tudja a komplementer szálat de novo megszintetizálni Ehhez kell egy primer, amely egy rövid oligonukleotid szakasz (RNS, DNS) komplementer a templát DNS egy szakaszával Az enzim az új bázisokat a primer 3’ OH végéhez adja hozzá, azaz az új szálat 5’-3’ irányban szintetizálja A primer eltűnik (kikerül) a szintetizálódó szálból Néhány állati vírus és a 29 bakteriofág RNS primer nélkül működik Mivel a szintézis az 5’->3’ irányba halad a hélix két szála nem replikálódhat egyszerre
A hélix kinyílik, kilakul a replikációs villa • Az egyik szál, vezér (leading) szál replikálódik • A komplementer szál a lagging strand, visszamaradt szál • Mivel a másik szálon is 5’->3’ a szintézis irány ott csak megszakításokkal mehet a szintézis • A fragmenseket felfedezőjükről Okazaki fragmenseknek nevezik
Replikáció iniciáció (indítás) Elongáció (a szál szintézise) Termináció (a replikáció befejezése) A termék(ek) megoldása A DNS replikáció lépései
A coli esetében a replikáció az oriC-nél kezdődik, kromoszóma replikációs origó (kezdőpont) • két iniciációs esemény is történik • mindkét replikációs villának van vezér és lagging szála
A replikációban több enzim is részt vesz E.coli-ban a DNS polimeráz III és polimeráz I A DNS Pol III a nagymennyiségű szintézisért felel Gyors működés, 60000 nukleotid/perc 5’->3’ polimeráz aktivitás Proofreading aktivitás 3’->5’ exonukleáz Az enzim magja három alegységből áll: ’ A DNS Pol I lassabb működésű Főleg a repair mechanizmusban játszik szerepet 5’->3’ polimeráz aktivitás 3’->5’ exonukleáz 5’->3’ exonukleáz aktivitás (primer) 5’ végéhez kapcsolódik, kivágja a ribonukleotidot, helyére dezoxiribonukleotidot épít, a primer elbomlik
Replikáció indítása (iniciáció) A DNS replikáció a kromoszóma replikációs origójánál kezdődik (oriC) A baktérium kromoszómának egy origója van Az élesztőnek több, ún. autonóm replikálódó szekvencia elem ~ 36 kb-onként található A baktériumokban az oriC konzervált struktúra és hasonlóság van a DnaA fehérjék között is
A DnaA fehérje ATP-t köt • 20-40 ilyen komplex az oriC 9 bp hosszúságú kötőhelyekhez kapcsolódik • Nukleoszómához hasonló szerkezet jön létre • Ha a DnaA fehérje egy a nem kódoló szálon lévő helyhez köt, akkor gátolja a transzkripciót azon a helyen • Az iniciációhoz a spirált szét kell csavarni (a HU fehérje és DNS giráz hatása is fontos) • Az oriC-hez közel eső erős promóterről (mioC) transzkripció indul, amely szintén segít a hélixet kitekerni • A DnaA fehérje köt az egyik DNS szálhoz • A helikáz loading faktorának (DnaC) segítségével a helikáz (DnaB) a DNS-hez kapcsolódik, majd ATP hidrolízis mellett a DnaC fehérje disszociál
A kitekert DNS-hez primer szintetizáló enzim kapcsolódik (primáz) Ez a komplex a repliszóma (primoszóma), amely a sejt egy részéhez kapcsolódik A komplex helyben marad, a DNS csúszik, sliding clamp Bacillusnál az oriC a pólusok felé helyezkedik el • a primáz mindkét szál esetén megszintetizálja a primert • Primáz helyett RNS polimeráz is szintetizálhatja a primert, de az hosszabb • A DnaA boxok között lévő GATC szekvencia metilált • A replikáció után egy ideig (kb. 10 perc) eltart míg az új szál metilálódik megakadályozza a további replikáció indítást • A korai iniciácót megakadályozza még • OriC nem kötődik a membránhoz a szegregáció miatt • DnaA/ATP komplexeknek is ki kell alakulniuk • Ugyanígy a szintetizálódó új szál nem rögtön metilálódik, ez lehetővé teszi a javítást
A DNS PolIII holoenzim komplex hozzákapcsolódik a primerhez, az új szál szintézise megindul A hélixet a helikáz kitekeri Az így keletkezett egyszálú részeket az SSB stabilizálja A komplex tau alegysége a helikázzal kapcsolódik össze ezáltal gyorsabb a helikáz előrehaladása A helikázt követi a komplex A DNS polimeráz csúszó bilincset alkot a DNS-en (két alegység) (ehhez előtte a clamp loader komplex szükséges és ATP hidrolízis) A DNS templát (minta) szál csúszik át a komplexen miközben szintetizálódik az új szál Az extra csavart, amit a helikáz alkotott a topoizomeráz oldja meg A natív szuperhelikális struktúrát a DNS giráz hozza létre A giráz es topoizomeráz egyensúlya szükséges a normál szerkezethez Elongáció
A replikáció terminációja E. coli-ban két terminációs lókusz ~180º-ra az oriC-től A két régió távolsága 350 kb, ellentétes irányultságú A két terminációs hely megakadályozza a replikációs villa tovahaladását az egyik rányban Bármelyik replikációs villa érkezzék is előbb, be kell várnia a másikat Így kiküszöbölhető az időzítés Nem ismert pontosan, hogyan terminálódik, de kell hozzá egy Tus fehérje A terminációs szekvencia coliban kettő 29 bp-os ismétlődő rész (inverted repeat) Bacillussubtilis-ben egy terC nevű terminációs lókusz van, 59 bp hosszú 2 fordítottan ismétlődő részből áll
A termékek megoldása A keletkezett DNS végeinek összekapcsolásakor három eset Megfelelő szál Katenát keletkezik, mint egy lánc Két független DNS molekula Ellentétes szál Konkatemer (folytonos, 2 vagy több egységnyi DNS molekula Megoldás Rekombináció, vagy topoizmeráz
DNS replikáció és osztódás összefüggése Baktérium haploid Genom ekvivalens Baktérium gyorsabban osztódik, mint a DNS-e replikálódik, mielőtt még befejeződne az egyik replikáció elkezdődik az újabb replikáció A genom a nukleotid bázispárok száma a kromoszómában Tobb genom ekvivalens is lehet gyorsan osztódó baktériumban Pl. egy baktérium, amiben félig replikálódott a genom 1,5 genom ekvivalens DNS-t tartalmaz Ettől még haploid, mert ugyanaz a DNS Ha kap külső DNS-t akkor ugyanabban citoplazmában két készlet is lehet abból a génből Ez a részleges diploidia (merodiploid) A populációban fennmaradhat sokáig, ha az plazmid, de ha nincs szelekciós előny, akkor eltűnik
Baktérium kromoszóma replikáció szabályozása E. coli gazdag táptalajban 20 perc alatt osztódik DNS replikáció 40 perc Megoldás (dichotóm) korai (éretlen) replikáció Gyors szaporodás esetén a replikáció vége előtt újabb iniciáció történik Ehhez szoros szabályozás kell A DnaA fehérje szintje meghatározó Az egy helyről (OriC) történő és korai (dichotóm) replikáció következménye, hogy az OriC közeli gének nagyobb dózisban lesznek jelen Ezért az oriC közelében olyan gének vannak, amelyek nagy mennyiségben szükségesek DnaA ez a szabályozáshoz szükséges Pl. 7 riboszomális RNS kódoló gén (amely az új sejt felépítéséhez szükséges)
Theta replikáció (kromoszóma, plazmidok) Rolling circle replikáció (plazmidok, fágok) Looped rolling circle
M13 Egyszálú DNS az érett fágban, dsDNS a replikatív forma
T7 fág Terminális redundancia