The trp operon
The trp operon. Reading: MVA pp. 760-767 and 1011-1018 Problem Ch 26: 9 Alberts: pp. 395-399. MVA Fig.21.1. MVA Fig.21.12. MVA Fig.21.13. MVA Fig.21.14. MVA Fig.21.15.


The trp operon
E N D
Presentation Transcript
Reading: MVA pp. 760-767 and 1011-1018 Problem Ch 26: 9 Alberts: pp. 395-399
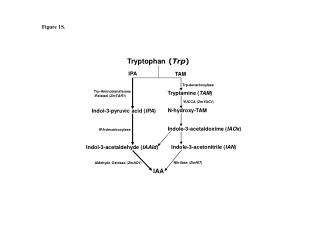
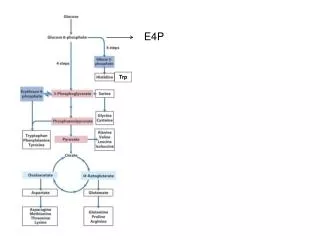
The chromosomal order of genes in the trp operon of E. coli and the sequence of reactions catalyzed by the enzyme products of the trp structural genes. The products of genes trpD and trpE form a complex that catalyzes specific steps, as do the products of genes trpB and trpA. Tryptophan synthetase is a tetrameric enzyme formed by the products of trpB and trpA. It catalyzes a two-step process leading to the formation of tryptophan. (PRPP, phosphoribosyl pyrophosphate; CDRP, 1-(o-carboxyphenylamino)-1-deoxyribulose 5-phosphate.) (After S. Tanemura and R. H. Bauerle, Genetics 95, 1980, 545.)
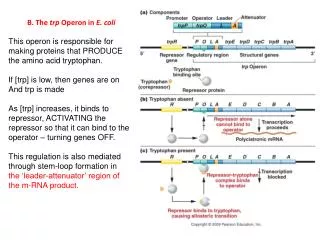
Originally, regulation of the trp operon was thought to occur solely through the repressor-operator system until deletion mutants located downstream of trpO were identified. These mutants displayed increased expression of the operon by six-fold which indicated the presence of an additional transcriptional control element. Why is repression not the only mode of regulation?
Evidence: • 1. Trp-tRNA synthetase mutants had regulatory anomalies. • 2. Addition of trp to trp-starved cells not only shut down • initiation of transcription but also inhibited transcription • already in progress on the initial segment of the operon. • 3. Mutants lacking a functional repressor could still respond • to trp starvation by increasing transcription of trp mRNA. • 4 . Deletion mutants in which both of the deletion termini were • within the transcribed region of the operon had an • unexpected six-fold increase in expression of the remaining • genes in the operon. Obviously, repressor binding was • unaffected.
5. Within the population of mRNAs produced in vivo from the 5' end of the trp operon, RNAs corresponding to the first 140 bp (the leader sequence) of the operon were several times more abundant than those from more distal regions, therefore a transcription termination site was located before the structural genes. 6. Starving bacteria of trp reduced termination at this site (the trp attenuator). 7. Mutations altering trp-tRNA synthetase, tRNAtrp or a tRNA trp modifying enzyme were found to decrease transcription termination at the trp attenuator. What does this suggest about the mode of attenuation?
8. Ribosome binding experiments with the 140 base transcript • demonstrated that ribosomes protect a 20 base segment from • nuclease attack. A potential AUG start codon is located in the • center of this region. • 9. A 14 residue peptide (the leader peptide) could be • synthesized from this start codon and contained tandem trp • residues near its C-terminus. • 10. The trp leader ribosome binding site was shown to be an • efficient site for the initiation of translation by fusing the leader • to a structural gene and demonstrating synthesis of the fused • polypeptide.
11. Two classes of termination defective leader mutants have been isolated. One type terminates at less than normal frequency and has bp changes in the 3:4 bp region. In vivo, these mutants have a 2-4 fold increase in operon expression. • 12. The second class of mutants have increased termination of • the attenuator. These prevent the relief from termination that • is associated with trp starvation. One of these mutants has an • altered start codon for the leader peptide. Another has a • G to A conversion at position 75, which would prevent • 2:3 pairing and cause formation of a 3:4 termination structure.
Genetic analysis indicated that the new control element was located in trpL, a 162 nt region 30-60 nt upstream from trpE. When trp is scarce, the entire 6720 nt polycistronic trp, including trpL, is synthesized. As the trp concentration increases, the rate of trp transcription decreases as a result of the trp r epressor-corepressor's greater abundance. With increasing [trp], the mRNA synthesized consists more and more of a 140 nt segment corresponding to trpL sequences only. The availability of trp results in the premature termination of transcription of the operon.