Flexibility of a polypeptide chain
190 likes | 496 Vues
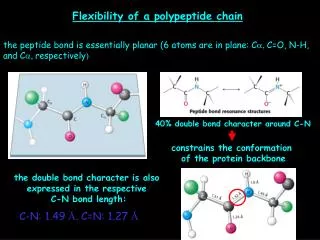
Flexibility of a polypeptide chain. the peptide bond is essentially planar (6 atoms are in plane: C a , C=O, N-H, and C a, respectively ). 40% double bond character around C-N. constrains the conformation of the protein backbone. the double bond character is also

Flexibility of a polypeptide chain
E N D
Presentation Transcript
Flexibility of a polypeptide chain the peptide bond is essentially planar (6 atoms are in plane:Ca, C=O, N-H, andCa, respectively) 40% double bond character around C-N constrains the conformation of the protein backbone the double bond character is also expressed in the respective C-N bond length: C-N: 1.49 Å, C=N: 1.27 Å
Protein conformation phi psi torsion (or dihedral) angles HN-CaandCa-CO are pure single bonds, high degree of freedom for rotation around these bonds lots of ways for a protein to fold if each amino acid has two main chain bonds to rotate about BUT! Any combination of phi and psi is possible?
Ramachandran plot many combinations are disfavored due to steric collisions (green regions are allowed,white regions are not)
What does thermodynamics have to say about protein folding? each copy of an unfolded polymer exists in a different conformation (ran- dom coil) yielding a mixture of many possible conformations, which would theoretically oppose folding due to favorable enthropy but in proteins, rigidity of peptide units and the restricted set of allowed phi and psi values actually limit the number of possible structures and it is overcome by interactions that favor the folded form e.g. hydro- phobic interactions among apolar side-chains (rather than being exposed to polar water), H-bonding network, S-S bridges, ion pairs, etc. a highly flexible polymer, of any kind, with large number of possibleconformations to adopt would NEVER fold into a unique 3D structure!!
Secondary structural elements alpha helix beta pleated sheet beta turn omega loop all forms stabilize via H-bridges between amino acids nearby in linear sequence alpha helix 3.6 residues/turn 100o rotation/residue rise/residue: 1.5 Å pitch of helix: 5.4 Å
Screw sense of an alpha helix right-handed (clockwise) or left-handed (counterclockwise) more stable, less steric clashes between side chains and backbone
helical content of a protein may vary from 0-100% Ferritin (iron storage protein) contains 75% alpha helix ~25% of all soluble proteins are largely helical an alpha helix is usually smaller than 45 Å proteins embedded in or crossing biological membranes build also up mainly from alpha helices
Beta (pleated) sheets (beta because this structure was the 2nd one, after the alpha helix, that Linus Pauling and Robert Corey envisioned/proposed in 1951, 6 years before the first ever protein structure determined by X-ray crystallography by Kendrew in 1957, myoglobin ) composed of 2 or more beta strands (fully extended chains) stabilized by H-bonding between polypeptide chains
purely parallel, purely antiparallel or mixed beta sheets exist 4-5 but even 10 or more strands make up a beta sheet beta sheets generally adopt a twisted shape: fatty acid binding proteins (important in lipid metabolism) almost entirely are built from beta sheets: MUSCLE FATTY ACID BINDING PROTEIN (1FTP.pdb)
Loops and turns globular proteins can be made up if turns and loops are incor- porated in structure beta turn = reverse turn = hairpin turn loop = omega loop (generally rigid, well-defined structures) loops and turns generally lie on the protein surfaces and par- ticipate in protein-protein and other types of interactions turn loops
Superhelices a-keratin (main component of wool and hair) consists of two right-handed a-helices intertwined to form a left-handed superhelix called a coiled coil (superfamily of coiled-coil proteins, ~60 proteins in humans) 2 or more a helices can entwine and form a stable, even 1000 Å (0.1 mm) or longer, structure found in cytoskeleton, filaments, muscle proteins 3.5 residues/turn, heptad repeats, every 7th residue is Leu on each strand and these two Leu interact (hydrophobic interaction), 2 Cys can also interact (S-S) stabilizing fiber wool can be stretched (some interactions among helices brake, S-S does not and pulls back after release) hair and wool have fewer cross-links, horn, claw, hoof are hard
Collagen most abundant protein in mammals, main fibrous component of skin, bone, teeth, cartilage and tendon extracellular protein, rod shape, ~3000 Å long/15 Å in diameter, 3 helical protein chains (~1000 residues each, every 3rd residue is Gly, Gly-Pro-(Pro-OH) triad is frequent, Pro-OH (4-hydroxyproline) is a natural amino acid derivative) no H-bonds inside the helical strands, stabilization occurs via steric repulsion between Pro and Pro-OH ~3 residues/turn, 3 helices wind in a superhelical cable that is stabilized by H-bond in between strands (Pro-OH participates in H-bonding network and lack of –OH on Pro in collagen lead to the disease scurvy (Vitamin C deficiency, ascorbate reduces Fe3+ to Fe2+ in prolyl hydroxylase for its continuous activity) Pro rings are on the outside, Gly in every 3rd position is needed because the superhelix is very crowded inside and there is no place for any other bigger amino acid
Tertiary structure the very first protein to be seen in atomic detail was myoglobin, the O2- carrier protein in muscle, determined by Kendrew in 1957 (6 Å resolution) Kendrew's original model of the myoglobin molecule, 1957, made of plasticine. single polypeptide chain, 153 residues, heme prosthetic (helper) group [heme: protoporphyrin IX and central iron ion], very compact molecule (45 X 35 X 25 Å), 70% of amino acids are in 8 a helices, the rest are in loops and turns
Heme hydrophobic amino acids are yellow, charged ones are blue, others are white cross-section
interior of globular proteins are rich in hydrophobic amino acids like Leu, • Val, Met, Phe • charged and rather polar residues, like Glu, Asp, Lys, Arg (Gln, Asn) localize • on the exterior of proteins • in myoglobin there are two critical His in the interior that conduct binding • of O2 • helices and b sheets may often have an amphipathic character: one part points towards the hydrophobic interior core of the protein, the other side points into solution burying polar main chain atoms in the hydrophobic interior is possible if all N-H and C=O moieties are in a H-bonding network (a helix, b sheet) proteins spanning biological membranes are the “exceptions that prove the rule” as they have a reverse distribution of hydrophobic and hydrophilic amino acids, like in porins, found in the outer membranes of bacteria (they are “inside out” relative to proteins function in aqueous environment)
Motifs and supersecondary structures certain combinations of secondary structure are present in many proteins and frequently exhibit similar functions, these combinations are called motifs or supersecondary structures For instance, a helix-turn-helix motif, often found in DNA-binding proteins some polypeptide chains fold into 2 or more compact globular units or regions that are connected by flexible regions, these are called domains (30-400 amino acids long) cell-surface protein CD4 consists of 4 similar domains
Quaternary structure proteins containing more than one polypeptide chains adopt a quaternary structure which describes the spatial arrangement of the subunits and the interaction between them each polypeptide chain is called a subunit the number of subunits may vary and we designate this by calling the protein a dimer, trimer, tetramer, etc. there can be homo- and hetero-multimers which may be tightened together covalently or non-covalently the Cro protein of bacteriophage l is a dimer of identical subunits
human hemoglobin , the O2-carrying protein of blood, consists of two a-type and two b-type subunits, a a2b2 hetero-tetramer viruses make the most out of limited genetic information: they have a protein coat that uses many, often identical, subunits repetitively in a symmetric array for their build-up: e.g. the rhinovirus, the cause of the common cold, includes 60 copies of each of four subunits forming a nearly spherical shell that encloses the viral genome electron micrograph of virus particles schematic view