Download
1 / 1
10 likes | 119 Vues
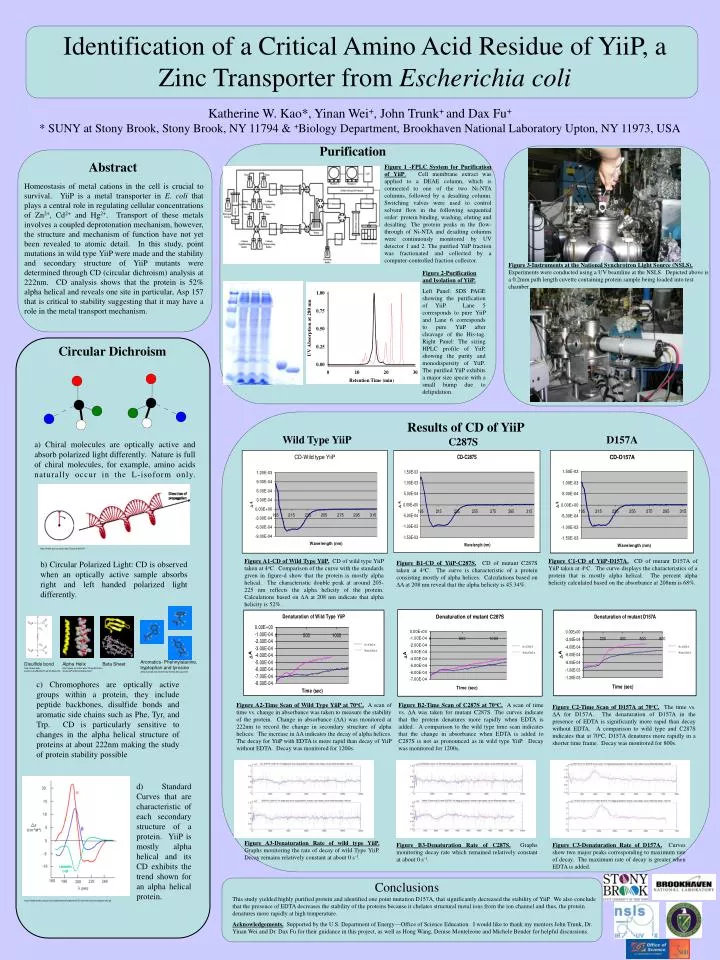
Identification of a Critical Amino Acid Residue of YiiP, a Zinc Transporter from Escherichia coli. Katherine W. Kao*, Yinan Wei + , John Trunk + and Dax Fu + * SUNY at Stony Brook, Stony Brook, NY 11794 & + Biology Department, Brookhaven National Laboratory Upton, NY 11973, USA. Purification.

E N D
Identification of a Critical Amino Acid Residue of YiiP, a Zinc Transporter from Escherichia coli Katherine W. Kao*, Yinan Wei+, John Trunk+ and Dax Fu+ * SUNY at Stony Brook, Stony Brook, NY 11794 & +Biology Department, Brookhaven National Laboratory Upton, NY 11973, USA Purification Abstract Figure 1 -FPLC System for Purification of YiiP. Cell membrane extract was applied to a DEAE column, which is connected to one of the two Ni-NTA columns, followed by a desalting column. Switching valves were used to control solvent flow in the following sequential order: protein binding, washing, eluting and desalting. The protein peaks in the flow-through of Ni-NTA and desalting columns were continuously monitored by UV detector 1 and 2. The purified YiiP fraction was fractionated and collected by a computer-controlled fraction collector. Homeostasis of metal cations in the cell is crucial to survival. YiiP is a metal transporter in E. coli that plays a central role in regulating cellular concentrations of Zn2+, Cd2+ and Hg2+. Transport of these metals involves a coupled deprotonation mechanism, however, the structure and mechanism of function have not yet been revealed to atomic detail. In this study, point mutations in wild type YiiP were made and the stability and secondary structure of YiiP mutants were determined through CD (circular dichroism) analysis at 222nm. CD analysis shows that the protein is 52% alpha helical and reveals one site in particular, Asp 157 that is critical to stability suggesting that it may have a role in the metal transport mechanism. Figure 3-Instruments at the National Synchrotron Light Source (NSLS). Experiments were conducted using a UV beamline at the NSLS. Depicted above is a 0.2mm path length cuvette containing protein sample being loaded into test chamber. Figure 2-Purification and Isolation of YiiP. Left Panel: SDS PAGE showing the purification of YiiP. Lane 5 corresponds to pure YiiP and Lane 6 corresponds to pure YiiP after cleavage of the His-tag. Right Panel: The sizing HPLC profile of YiiP, showing the purity and monodispersity of YiiP. The purified YiiP exhibits a major size specie with a small bump due to delipidation. Circular Dichroism Results of CD of YiiP Wild Type YiiP D157A C287S a) Chiral molecules are optically active and absorb polarized light differently. Nature is full of chiral molecules, for example, amino acids naturally occur in the L-isoform only. http://www.sinc.sunysb.edu/Class/che321ff/ Figure A1-CD of Wild Type YiiP. CD of wild type YiiP taken at 4oC. Comparison of the curve with the standards given in figure-d show that the protein is mostly alpha helical. The characteristic double peak at around 205-225 nm reflects the alpha helicity of the protein. Calculations based on DA at 208 nm indicate that alpha helicity is 52%. Figure C1-CD of YiiP-D157A. CD of mutant D157A of YiiP taken at 4oC. The curve displays the characteristics of a protein that is mostly alpha helical. The percent alpha helicity calculated based on the absorbance at 208nm is 68%. b) Circular Polarized Light: CD is observed when an optically active sample absorbs right and left handed polarized light differently. Figure B1-CD of YiiP-C287S.CD of mutant C287S taken at 4oC. The curve is characteristic of a protein consisting mostly of alpha helices. Calculations based on DA at 208 nm reveal that the alpha helicity is 45.34%. No EDTA No EDTA No EDTA With EDTA With EDTA With EDTA Aromatics- Phehnylalanine, tryptophan and tyrosine www.sunysb.edu/chemistry/molecules/aa.html Disulfide bond http://www.web-books.com/MoBio/Free/Ch2A3.htm Alpha Helix http://www.sci.sdsu.edu/TFrey/ProtStructClass/ProtStructClass.html Beta Sheet c) Chromophores are optically active groups within a protein, they include peptide backbones, disulfide bonds and aromatic side chains such as Phe, Tyr, and Trp. CD is particularly sensitive to changes in the alpha helical structure of proteins at about 222nm making the study of protein stability possible Figure A2-Time Scan of Wild Type YiiP at 70oC. A scan of time vs. change in absorbance was taken to measure the stability of the protein. Change in absorbance (DA) was monitored at 222nm to record the change in secondary structure of alpha helices. The increase in DA indicates the decay of alpha helices. The decay for YiiP with EDTA is more rapid than decay of YiiP without EDTA. Decay was monitored for 1200s. Figure B2-Time Scan of C287S at 70oC. A scan of time vs. DA was taken for mutant C287S. The curves indicate that the protein denatures more rapidly when EDTA is added. A comparison to the wild type time scan indicates that the change in absorbance when EDTA is added to C287S is not as pronounced as in wild type YiiP. Decay was monitored for 1200s. Figure C2-Time Scan of D157A at 70oC. The time vs. DA for D157A. The denaturation of D157A in the presence of EDTA is significantly more rapid than decay without EDTA. A comparison to wild type and C287S indicates that at 70oC, D157A denatures more rapidly in a shorter time frame. Decay was monitored for 800s. d) Standard Curves that are characteristic of each secondary structure of a protein. YiiP is mostly alpha helical and its CD exhibits the trend shown for an alpha helical protein. Figure A3-Denaturation Rate of wild type YiiP. Graphs monitoring the rate of decay of wild Type YiiP. Decay remains relatively constant at about 0 s-1. Figure B3-Denaturation Rate of C287S. Graphs monitoring decay rate which remained relatively constant at about 0 s-1. Figure C3-Denaturation Rate of D157A. Curves show two major peaks corresponding to maximum rate of decay. The maximum rate of decay is greater when EDTA is added. Conclusions This study yielded highly purified protein and identified one point mutation D157A, that significantly decreased the stability of YiiP. We also conclude that the presence of EDTA decreases the stability of the proteins because it chelates structural metal ions from the ion channel and thus, the protein denatures more rapidly at high temperature. Acknowledgements. Supported by the U.S. Department of Energy—Office of Science Education. I would like to thank my mentors John Trunk, Dr. Yinan Wei and Dr. Dax Fu for their guidance in this project, as well as Hong Wang, Denise Monteleone and Michele Bender for helpful discussions. http://employees.csbsju.edu/hjakubowski/classes/ch331/protstructure/cdsecondst.gif