Chapter 6: Enzymes
Chapter 6: Enzymes. From Wikipedia: Enzymes are proteins that catalyze (i.e., increase the rates of ) chemical reactions. Nearly all known enzymes are proteins. However, certain RNA molecules can be effective biocatalysts too. These RNA molecules have come to be known as ribozymes.

Chapter 6: Enzymes
E N D
Presentation Transcript
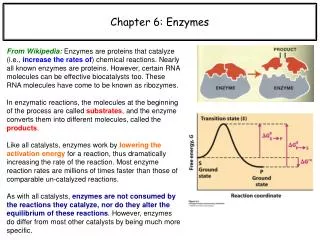
Chapter 6: Enzymes From Wikipedia: Enzymes are proteins that catalyze (i.e., increase the rates of) chemical reactions. Nearly all known enzymes are proteins. However, certain RNA molecules can be effective biocatalysts too. These RNA molecules have come to be known as ribozymes. In enzymatic reactions, the molecules at the beginning of the process are called substrates, and the enzyme converts them into different molecules, called the products. Like all catalysts, enzymes work by lowering the activation energy for a reaction, thus dramatically increasing the rate of the reaction. Most enzyme reaction rates are millions of times faster than those of comparable un-catalyzed reactions. As with all catalysts, enzymes are not consumed by the reactions they catalyze, nor do they alter the equilibrium of these reactions. However, enzymes do differ from most other catalysts by being much more specific.
Enzymes work by lowering activation energy barriers k+1 k+2 k+3 k+1 E + S ES EP E + P k-1 k-2 k-3 k-1 Recall:The change in free energy for a reaction at equilibrium (DG°) is related to the equilibrium constant, which is Keq = [P]eq/[S]eq, by the following relation: DG° = -RT lnKeq. Both of the above quantities are thermodynamic measurements, meaning they are unaffected by the presence/absence of an enzyme. Enzymes only affect the activation energy (DG‡), which is related to the reaction kinetics via the Arrhenius equation: rate = A·exp(-DG‡/RT). S P
Binding energy vs. stabilization of the TS (it’s not as simple as your text implies)
Initial velocity (n) of an enzyme catalyzed reaction Recall:The rate of a reaction is concentration dependent… n = k[S]. However, the rate of [S] will decrease as the reaction proceeds, making the rate also decrease. We evaluate enzyme power by quantifying reaction rate enhancements. To eliminate the effects of decreasing [S], we only track the initial velocity (sometimes designated as no) of a reaction. Under this initial regime (the first 60-75 seconds), the change in [S] should only be a few percent, and thus acceptable. As shown to the left, after some time, the effect of decreasing [S] drastically affects the reaction rate.
Michelis-Menten kinetics k+2 k+1 E + S ES E + P k+2 k+3 k-1 k+1 E + S ES EP E + P k-2 k-3 k-1
Lineweaver-Burke (the double reciprocal plot)
Interpreting Km Km is commonly interpreted as the relative enzyme affinity for its substrate. This interpretation is acceptable, if and only if, product formation is rate limiting (meaning, k2 << k-1). However, if product formation is not clearly rate limiting, then Km is actually an effective rate constant, describing multiple steps. Note: the blue line could actually cross more than just 2 barriers. For example: E + S ES ES’ ES’’ EP E + P
Km values can vary widely, even for different substrates to the same enzyme
Interpreting kcat (the turnover number) Consider the situation where product release is limiting: ES E + P (refer back to the purple box in slide #11). It is useful to define a more general rate constant to describe this situation. When product release is limiting, kcat is equivalent to the number of substrate molecules converted to product in a given unit of time (thus, turnover number). However, like Km, kcat is generally a complex function of multiple rate limiting steps. As with Km, kcat can vary a lot…
The only generally useful measure of catalytic power is enzyme efficiency: kcat / Km NeitherKm and kcatare sufficient to describe ‘how good an enzyme is’ because their simplistic interpretations are only useful under fairly narrow assumptions. Note that in cellular conditions, [E] and [S] << Km… Meaning kcat / Km is an effective 2nd order rate constant that measures how efficiently an enzyme converts substrate to product. The most efficient enzymes are diffusion limited (108-1010 M-1s-1), meaning the slow step is bringing the substrate to the enzyme. These enzymes are termed perfect enzymes. Either a large value of kcat (rapid turnover) or a small value of KM (high affinity for substrate) makes the ratio kcat / Km large...
Multiple substrates Ternary complex Ping-pong mechanism
CuZnSOD a perfect enzyme with a ping-pong mechanism E-Cu+++ O2-→E-Cu++ O2 E-Cu++ O2- + 2H+→E-Cu+++ H2O2 2O2- + 2H+→ O2 + H2O2 Ping-pong mechanism
Electrostatics make CuZnSOD “better than perfect” Cationic funnel of CuZnSOD Copper ion
Electrostatic potential maps across the CuZnSOD protein family are evolutionarily conserved REF: Livesay et al., Biochemistry, 2003, 42(12):3464-3473.
Substrate binding: induced fit vs. lock-and-key Apo-hexokinase Holo-hexokinase
Competitive inhibitors lower the enzyme’s effective affinity (Km) for the substrate vs.
Noncompetitive inhibitors lower the effective Vmax, but have no affect on Km (this is not in your text) Noncompetitive inhibition is a type of enzyme inhibition that reduces the maximum rate of a chemical reaction (Vmax) without changing the apparent binding affinity of the catalyst for the substrate (Km). Noncompetitive inhibition usually applies to enzymes and differs from competitive inhibition in that the inhibitor always binds to the enzyme at a site other than the enzyme's active site (this other site is called an allosteric site). A noncompetitive inhibitor reduces the effective enzyme concentration [Eeff] = [Etot] – [INC]. vs.
Uncompetitive inhibitors affect both Vmax and Km Uncompetitive inhibition takes place when an enzyme inhibitor binds only to the ES. This reduction in the effective [ES] increases the enzyme's apparent affinity for the substrate through Le Chatelier's principle (Km is lowered) and decreases the maximum enzyme activity (Vmax), as it takes longer for the substrate or product to leave the active site. Uncompetitive inhibition works best when substrate concentration is high.
Mixed inhibitors bind to both E and ES, and affect both Km and Vmax Mixed inhibition refers to a combination of two different types of reversible enzyme inhibition – competitive inhibition and uncompetitive inhibition. The term 'mixed' is used when the inhibitor can bind to either the free enzyme or the ES complex. In mixed inhibition, inhibitor binding alters the ratio of Km / Vmax, which is constant in uncompetitive inhibition. Note: noncompetitive inhibition is really just a (hypothetical) subset of mixed inhibition where the change in Km / Vmax is only occurring within the effective Vmax.
Summary of inhibitor effects (note that I have added the last line) Noncompetitive Vmax / a’ Km (b/c a= a’)
The effect of pH Q: Can you predict the effect of temperature on no?
You will need to know three enzyme mechanisms… (1.) Alcohol Dehydrogenase (2.) Enolase (3.) Chymotrypsin
Dehydrogenases catalyze RedOx reactions Oxidized (NAD+) Reduced (NADH)
Alcohol dehydrogenase ADH requires both NAD+ and Zn+2 ADH Alcohol + NAD+ Acetaldehyde + NADH
Alcohol dehydrogenase: a simple mechanism ADH stabilizes the oxyanion intermediate!
Enolate intermediate The enolate anion is formed by loss of a proton from the a-carbon of a carbonyl. For example, consider acetone… The enolate anion is stabilized by resonance
ChymotrypsinogenChymotrypsin Chymotrypsinogen is a precursor of the digestive enzyme chymotrypsin (zymogen). This molecule is inactive and must be cleaved by trypsin, and then by other chymotrypsin molecules before it can reach its full activity. The active site of the chymotrypsinogen is covered by a six-amino-acid-long mask. It is only when this mask is removed - when it enters the lumen of the intestine and comes into contact with chymotrypsin molecules - that the enzyme becomes active. This is a very useful safety feature for a protein-digesting enzyme. If it were not inactivated in this way, it would digest the pancreas where it is produced.
Chymotrypsin is a serine protease enzyme, meaning it ‘chops up’ other proteins
Chymotrypsin: a fairly complicated multi-step mechanism From Wikipedia: An example of a catalytic triad is present in chymotrypsin, where the triad consists of S195, D102, and H57. In essence, S195 binds to the substrate polypeptide to the side of a phenylalanine, tryptophan, or tyrosine residue closer to the C-terminus, holding it in place. D102 and H57 then hydrolyze the bond. This takes place in several steps. Upon substrate binding, the D102 carboxylate hydrogen bonds to H57, increasing the pKa of its imidazole nitrogen from 7 to about 12, making it a powerful general base that deprotonates S195. The deprotonated S195 serves as a nucleophile, attacking the carbonyl carbon on the C-terminal side of the residue, which forces the carbonyl oxygen to accept an electron, and transforming the sp2 carbon into a tetrahedral intermediate. This intermediate is stabilized by an oxanion hole, which also involves S195. 3. Collapse of this intermediate back to a carbonyl causes H57 to donate its proton to the nitrogen attached to the alpha carbon. The nitrogen and the attached peptide fragment leave by diffusion. 4. A water molecule then donates a proton to H57 and the remaining --OH- attacks the carbonyl carbon, forming another tetrahedral intermediate. The OH is a poorer leaving group than the C-terminal fragment, so when the tetrahedral intermediate collapses again, S195 leaves and regains a proton from H57. 5. The cleaved peptide, now with a carboxyl end, leaves by diffusion.
Oxyanion holes stabilize the tetrahedral oxyanion intermediate From Wikipedia: It was discovered that additional amino acids of chymotrypsin, Gly193 and Ser195, are involved in creating what is called an oxyanion hole. Both donate backbone hydrogens for hydrogen bonding. When the tetrahedral intermediates are generated, the negative oxygen ion, having accepted the electrons from the carbonyl double bond fits perfectly into the oxyanion hole. In effect, serine proteases preferentially bind the transition state and the overall structure is favored, lowering the activation energy of the reaction. This "preferential binding" is responsible for much of the catalytic efficiency of the enzyme. Oxyanion holes are common catalytic motifs (e.g., ribose-5-phosphate isomerase) Serine proteases oxyanion hole
The oxyanion hole in triosephosphate isomerase is constructed from cationic sidechains Another example, triosephosphate isomerase: