Download
1 / 22
240 likes | 770 Vues
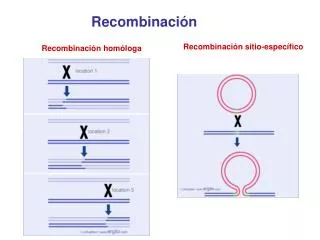
Recombinación. Recombinación sitio-específico. Recombinación homóloga. Enzimas involucradas en la generación de la cadena sencilla con el extremo 3´OH libre. sitio chi 5´GCTGGTGG3´ 3´CGACCACC5´. RecBCD helicasa y nucleasa que forman el sustrato para RecA. Filamento de Rec A. Rec A.

E N D
Recombinación Recombinación sitio-específico Recombinación homóloga
Enzimas involucradas en la generación de la cadena sencilla con el extremo 3´OH libre sitio chi 5´GCTGGTGG3´ 3´CGACCACC5´ RecBCD helicasa y nucleasa que forman el sustrato para RecA
Resolución de las uniones Holliday Ruv A se une a las cuatro hebras del intermediario de Holliday Ruv B es hexámero con actividad de ATPasa que sirve de motor para su movimiento. El consumo de ATP permite girar a la molécula Ruv C es una endonucleasa que resuelve los intermediarios de Holliday En eucariontes no se han encontrado homólogos para las proteínas Ruv, pero sí para la Rec A
Resolución de las uniones Holliday La única diferencia entre los dos cromosomas En algunos casos, la doble hélice de abajo rotará 180 o. Este proceso se llama Isomerización. resolucion isomerización
Reparación del ADN Enfermedades humanas asociadas a problemas en los mecanismos de reparación
Depurinación y desaminación de bases Sitios AP 5000 al día 100 al día
La naturaleza de las bases permite detectar mejor el daño
Dímeros de timina Exposición a radiaciones ultravioleta
Daños que pueden producir los errores en el DNA si no se reparan
MECANISMOS DE REPARACION • 1- Sistemas de Reparación directos • Enzimas que revierten directamente el daño. • Por estos mecanismos se reparan: metilación de guanina, y en algunos vertebrados dímeros de pirimidína. No intervienen nucleasas ni ADN-polimerasas. • Fotoreactivación(ruptura de los dímeros de pirimidinas por acción de una fotoliasa (phr) activada mediante luz visible). • 2- Sistemas de Reparación Indirecta • Hay intervención de nucleasas y ADN-polimerasas. Se necesita de la hebra “molde” perteneciente al mismo cromosoma o al homólogo. • Reparación por Escisión (BER , NER, MMR) • Reparación de nucleótidos (NER)aislados por lesión UV: necesita la otra hebra como templado (hasta 30 bp). Intervienen las endonucleasas uvrA,B,C y la helicasa uvrD. • Además de foto productos, repara lesiones voluminosas (bulky) que distorsionan la conformación del dúplex y que obstaculizarían la transcripción y replicación.
Reparación de bases modificadas (BER).- Repara casos de alteraciones puntuales en bases nitrogenadas (lesiones NO voluminosas) producidas por alquilación, oxidación o desaminación. Se origina un “sitio AP” y luego se retira el nucleótido “AP” y se re sintetiza la hebra) • Reparación post- replicativa • Reparación del apareamiento (MMR) (“mismatch repair”): • Su principal tarea es remover bases mal aparadas y pequeños “loops” introducidos por inserciones / deleciones durante la replicación • una metilasa reconoce DNA recientemente replicado (dam) e intervienenlas proteínas“mut”(helicasas, etc). En otros organismos pueden ser otras señales. • Reduce los errores de replicación de 10-7 a 10-10 pb / replicación
… Reparación post- replicativa • Recombinación Homóloga (HR) • Reparación de ambas cadenas • Usa ADN homólogo como templado y es altamente exacto • Más activo durante la Fase S y G2 • Unión de extremos no homólogos (NHEJ) • Reparación de ambas cadenas • No usa ADN templado y generalmente se pierden algunos nucleótidos. • Más activo en la Fase G1
1)Reparación Directa de Daño al DNA Fotoreactivación • Sistema de reparación activado por presencia de luz. • Fotoliasa.-Detecta al DNA dañado y se une a éste. La enzima absorbe luz azul y se activa. Rompe los enlaces covalentes entre los dímeros de timina. • La enzima se disocia y se separa del DNA.
Reparación por escisión de Bases (BER) 1) Iniciado por DNA glicosilasa específica reconoce el daño, corta la unión glicosílica entre base y azúcar y se forma el sitio AP 2) Sitio AP reconocido por AP endonucleasa (corte 5’ de AP). 3) Fosfodiesterasa (corte 3’). 4) DNA polimerasa rellena el gap DNApol I (E.coli), DNA pol (mamíferos). 5) DNA ligasa 2) Reparación Indrecta de Daño al DNA
Reparación por escisión de Nucleótidos (NER) Escinucleasa uvrABC realiza este tipo de reparación en dímeros de timina, otros fotoproductos y bases dañadas. Escinucleasa (246 kDa) está compuesta por tres subunidades (A, B y C) UvrAse une al DNA en la región dañada. UvrB/UvrC tienen actividad de endonucleasa y corta en los lados adyacentes de la cadena liberando un oligonucleótido La región “vacía” es rellenada por una DNA polimerasa I y sellada por una DNA ligasa. E.coliSistema Uvr ABC: Remoción de 12nt EucariontesRemoción de 24-29 nt
Reparación de bases mal apareadas(MMR) E.coli genes mut S, L, H Reemplaza hasta 1kb Metilación diferencial (dam, dcm) MutS reconoce el mismatch MutH distingue ambas cadenas Corte en GATC en la cadena no metilada Mut L coordina actividad de Mut S y H En eucariotas homólogos de proteínas Mut