Quasispecies Assembly Using Network Flows
Alex Zelikovsky Georgia State University Joint work with Kelly Westbrooks Georgia State University Irina Astrovskaya Georgia State University David Campo Centers for Disease Control Yury Khudyakov Centers for Disease Control Piotr Berman Pennsylvania State University


Quasispecies Assembly Using Network Flows
E N D
Presentation Transcript
Alex Zelikovsky Georgia State University Joint work with Kelly Westbrooks Georgia State University Irina AstrovskayaGeorgia State University David Campo Centers for Disease Control Yury Khudyakov Centers for Disease Control Piotr Berman Pennsylvania State University Ion Mandoiu University of Connecticut Quasispecies Assembly Using Network Flows
Outline • 454 Sequencing of Virus Genome • Quasispecies Assembly • The Read Graph • Network Flow Formulations • Phasing Flow Problem • Maximum Likelihood • Simulation Results
HCV Quasispecies • HCV is a small, enveloped, positive sense single strand RNA virus that is responsible for Hepatitis C infection. • Over the course of infection, the mutations made in replication are passed down to descendants, eventually producing a family of related variants of the ancestral genome referred as quasispecies. • Due to HCV's high mutation rate, in time the quasispecies in an infected person can become very diverse. • A better understanding of HCV quasispecies diversity could potentially lead to new treatments. • The ultimate objective of our work is to develop a method of computationally inferring the quasispecies sequences in a HCV-infected individual.
454 Sequencing • 454 Sequencing is one promising technology that may prove useful for quasispecies sequencing. It is a massively-parallel pyrosequencing system developed the by biotechnology firm 454 Life Sciences for genome sequencing. • The system fragments the source genetic material to be sequenced into pieces called reads. Then, each read is sequenced and the original genome is reconstructed via software. • Since this system was originally designed to sequence genetic material from a single organism, the software assembles all of the reads to a single genome. • In order to use 454 for quasispecies sequencing, new methods and software are needed to correctly infer the sequences of the quasispecies present in an infected person and their population frequencies directly from 454 read data.
Problem Formulations • Given a collection of 454 reads taken from a quasispecies population of unknown size and distribution, reconstruct the quasispecies population, i.e. the sequences and tehir frequencies. Original Quasispecies 454 Reads Reconstructed Quasispecies
Parsimonious/Min Cost Quasispecies Assembly • Given a set of reads, • Find the minimum number set of quasispecies covering all reads • Given a set of reads with costs on read overlaps, • Find the minimum number set of quasispecies covering all reads • The cost of the assembly should be inversely connected to the likelihood that the assembly is the correct one. Reconstruction 1 Cost: C1 Original Quasispecies 454 Reads Reconstruction 2 Cost: C2 If C1 < C2, then favor Reconstruction 1 over Reconstruction 2
Read Alignment • Before beginning assembly, first find the genome offset of the read. We assume that the consensus sequence for the particular strain of HCV that the quasispecies came from is available to us. • We simply align each read to the consensus sequence, choosing the offset that yields the best alignment (i.e. lowest Hamming distance). • Because HCV quasispecies don't contain repeats as long as a 454 read, the alignment is both fast and extremely accurate. GUCUCAUCGGAACAGCAAAACACUUGCCCCGAACGCUAGCGGUUGGGGUACUAUUCAAUGGCUGUAG AACAGCAAAACACUUGCUCCGAACGCUAGCGGUUGGGGAACUAU
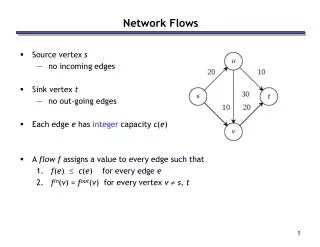
The Read Graph • The data structure: a directed acyclic graph that contains every possible quasispecies reconstruction. • An aligned read can be contained within another aligned read. • Find the subset of reads that are not contained within any other read • We call these reads “superreads” • “subreads” = everything else • The superreads are vertices in the read graph.
Edges of the Read Graph • Put an edge between read X and read Y if • X overlaps Y in the alignment • some suffix of X = some prefix of Y UGGACUAGAUGUGGUGGGUGCUCUCCGGAAUACCUUGGUGGCGGGU GAUGUGGUGGGUGCUCUCCGGAAUACCUUGGUGGCGGGUUAGAGA GGGUGCUCUCCGGAAUACCUUGGUGGCGGGUUAGAGAGAAGAGAGCA CUCCGGAAUACCUUGGUGGCGGGUUAGAGAGAAGAGAGCAAGUGUCA AUACCUUGGUGGCGGGUUAGAGAGAAGAGAGCAAGUGUCAACGCCUA
Quasispecies in the Read Graph • Then, we add two extra vertices: a new vertex with outgoing edges to all vertices with indegree 0 (the “source”) and a new vertex with incoming edges from all vertices with outdegree 0 (the “sink”) The Read Graph Source Sink Any path from the source to the sink represents a potential sequence in the quasispecies population!
Transitive Reduction • Edge u w logically follows from edges u v and v w • Drop edge u w from consideration – no information, any quasispecies sequence containing u and w will also have v • The transitive reduction of a graph • = smallest subgraph that maintains all reachability relationships • The graph is partially closed – the transitive reduction found in O(δ|E|), where δ is the read degree
Estimating Read Frequencies • In general, superreads may be contained in several quasispecies sequences. • Thus, each superread has associated with its frequency = the sum over the quasispecies of the population frequencies of quasispecies that contains the superread. • Although the true read frequencies are unknown to us, we may estimate them by counting the number of subreads contained within each superread. • By definition, the read frequency of the source and sink vertices are 0.
Probability of a True Overlap • Given N reads over Q sequences, each read with L possible starting positions, the probability that a position is bu for some read u is N/(LQ). • Let (u, v) be an edge in transitive reduction. • The probability of bv-bu > Δ is proportional to exp(Δ N/(LQ)). • Probability of an edge from the source or to sink is 0. bu GUGGGGGCAGCGGACGUAUGC GACGUAUGCAGAACUCUAGGCA Δ bv
Network Flow Through Vertices • Replacing the vertex for read r with • two vertices r_b and r_e • and the edge (r_b, r_e)
Networks Flows • Observe that the true quasispecies sequences in the read graph can be represented as a flow: 1 1 1 1 2 1 3 1 2 2 2 2 1 1 2 1 1 1 Each vertex has a frequency proportional to the number and frequencies of the quasispecies that contains it's associated superread. When we solve the flow, we demand that each vertex has a inflow passing into it >= its frequency.
Min Cost Flow • We define the cost of a flow in the following manner: • The flow cost of an edge is that edge's cost multiplied by the amount of flow that traverses the edge • The cost of a flow through the graph is the sum of all of the edge flow costs. • Out of all possible flows that go through all of the vertices in the graph, we seek to find the flow with the minimum cost. • After solving min cost flow for the graph, all of the edges that have flow > 0 are assumed to participate in true quasispecies. The remaining edges can be dropped from the graph.
LP for Min Cost Flow • Although there are fast combinatorial algorithms for solving min cost flow, we opted to solve the flow using a linear program. • For each edge e, create a real-valued, nonnegative variable fe to represent the flow across that edge. The Read Graph Source Sink
Linear Program for Min Cost Quasispecies Assembly • Objective:Minimize the sum of cost(e) * fe over all edges e in the read graph.Subject to: • For all vertices v: • The sum of fe over incoming edges to v equals the sum of fe over outgoing edges from v. • The sum of fe over incoming edges to v is greater or equal to the frequency of v.
Splitting Flow in Quasispecies • Five quasispecies share a common long segment [a,b] and differ on the left and the right in value of a SNP. The resulted graph with network flow have multiple feasible solutions. l a b r b-a > the read length A A C C T T T A CT Multi set L Multi set R A f=2 T f=3 f=2 f=1 C A f=1 T f=1 C
Quasispecies Matching Problem • Given • two multisets of haplotypes • L on the segment [l, b] and • R on the segment [a, r] • such that all haplotypes are indistinguishable on a common • segment [a, b] (l < a < b < r), |L| = |R|, • Find • the matching between multisets L and R such that • concatenation of the matched haplotypes • correspond to the original quasispecies.
Decomposing the flow into paths • General strategy: • Find a source-to-sink path with positive flow f • Subtract f from all of the edge flows in the path • How to find paths? • Pick the shortest path ⇒ “most likely quasispecies” • Pick the maximum bandwidth path ⇒ “most frequent quasispecies” 1→3→5→6 is the shortest path 1→2→4→6 is the maximum bandwidth path
Finding Max Bandwidth Paths • A single iteration of the Bellman-Ford algorithm gives an efficient method for finding the maximum bandwidth path from the source to the sink: • Initialize: • For each vertex I • W(i) ← 0 • p(i) ← 0 • For the source s • W(s) ← +infinity • Relax: • For each edge (i,j) in order *** (i,j)< (k,l) if i<k or i=k & j<l *** • if W(j) < min { W(i), cap(i,j) } *** cap(i,j) capacity of (i,j) • W(j) ← min { W(i), cap(i,j) } • p(j) ← I • Return path p(sink), p(p(sink)),..., source=0 • Finding minimum cost paths is simple: just grow a shortest-path tree from the source using costs for weights.
Maximum Likelihood Choice • After path decomposition, we have a set of candidate quasispecies sequences, but we don’t know what their frequencies are. • Given a set of candidate quasispecies and observed reads • Expectation Maximization: alternates between 2 steps until convergence: Expectation (E) and Maximization (M) • E Step: Calculates the expected likelihood by including the current estimate of the latent variables • M step: Computes maximum likelihood estimates of parameters by maximizing the expected likelihood found in the previous E step
EM Implementation • Create a bipartite graph: • Left side: quasispecies • Right side: superreads • Put an edge if quasispecies contains read • Keep 3 sets of numbers: • For each qsp q, keep its estimated frequency fq • For each superread r, keep its frequency nr • For each (q, r) edge, keep pqr • E step: Compute pqr = nr · fq / Σ fq for every edge • M step: Compute fq = Σ pqr / Σ nr for every qsp
Validation • Real quasispecies population, simulated reads • Real data: 44 sequences from E1E2 region of HCV • 3 populations consisting of 10 sequences each: • Uniformly distributed frequencies (the “uniform” population) • Geometrically distributed frequencies (the “geometric” population) • Highly skewed distribution (the “skewed” population)
Instance Generation • Inputs: A quasispecies population Q, n = number of reads desired n, read length mean μ and variance σ2 • S ←∅ • While |S| < number of reads desired • Randomly select a quasispecies q of length lq according to the population frequency distribution • Generate a read length lr using normal distribution (μ, σ2) • Generate an offset o using uniform distribution on [0, lq - lr] • Extract a substring of length lr starting at position o from quasispecies q and add it to S • Return: S
Quality Measures • Percentage of correctly predicted sequences: • Takes into account frequencies • Symmetric difference between multisets • “Switching error” • Generalization of “switching error” from the haplotype phasing community • Average number of times each path corresponding to a predicted quasispecies switches between paths in the read graph corresponding to real quasispecies.