

Транскрипція
E N D
Presentation Transcript
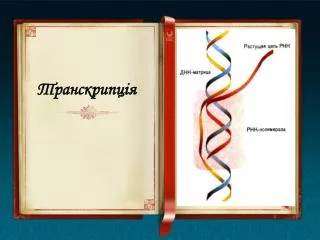
Транскрипція - це процес біосинтезу РНК на матриці ДНК. Копії мРНК, синтезовані в процесі транскрипції, переносяться в цитоплазму, на місце синтезу білка, де самі стають матрицями білкового синтезу. Таким чином, дуже важливо, щоби в процесі транкрипції копіювався не будь-який довільний відрізок ДНК, а змістовний. Така ділянка ДНК, яка несе інформацію про будову певного білка, або білків називається геном.
Одиницю процесу транскрипції називають опероном. В процесі транскрипції синтезуються крім мРНК також інші види РНК, необхідні для біосинтезу білків (тРНК, рРНК).
Фермент, який каталізує утворення копії РНК, - ДНК-залежна-РНК-полімераза - починає синтез в певній ділянці ДНК - промоторі, і закінчує його також в строго визначеній ділянці - термінальній. Цей фермент складається з чотирьох субодиниць і кислого білка -s-фактора. s-фактор забезпечує приєднання РНК-полімерази до ДНК саме в ділянці промотора, тобто узнавання місця початку транскрипції. Відомі різні -s-фактори, специфічні до промоторів різних наборів генів. Для синтезу необхідні всі чотири типи рибонуклеозидтрифосфатів (АТФ, ГТФ, ЦТФ і УТФ, а також йони Mg2+.
Оскільки РНК-полімераза може починати ланцюг з об’єднання двох мононуклеотидів, для транскрипції не потрібна затравка. РНК-полімераза, а саме її s-фактор, узнає ділянку ДНК - промотор. Встановлено, що на цій ділянці міститься специфічна послідовність нуклеотидів: (поліА і поліТ). Також цій ділянці властиві послідовності нуклеотидів з внутрішньою віссю симетрії, наприклад (по аналогії з паліндромами – текстами з внутрішньою віссю симетрії, наприклад “А роза упала на лапу Азора”) – АЦЦТГГЦТАГЦЦАГГТ, які можуть утворювати “шпильковидні вирости” полінуклеотидного ланцюгу, які служать сигнальними ділянками при узнаванні білками – ферментом і регуляторами.
Полімераза шукає цю ділянку серією випадкових асоціацій, тобто шляхом проб і помилок, що триває всього 15-20 секунд. Фермент приєднується до дволанцюгової матриці, викликаючи локальну денатурацію ДНК, і починає зчитування з ланцюга, який містить послідовність піримідинових нуклеотидів (ТТТТТТ) – кодуючий ланцюг. РНК-копія ідентична за виключенням заміни Т на У і пентози комплементарному, некодуючому ланцюгу ДНК.
Ініціація синтезу полягає у взаємодії субстратного центра ферменту з двома першими нуклеотидами копії, комплементарними матриці, і об’єднанні їх. Першим є пуриновий нуклеотид.
Елонгація, тобто нарощування полінуклеотидного ланцюга, забезпечується пересуванням РНК-полімерази по матриці, і здійснюється з швидкістю 40-50 нуклеотидів за секунду. По мірі просувння ферменту по матриці відбувається розплітання ДНК. Розмірн розплетеної ділянки становить біля 17 нуклеотидних пар.
В ділянці термінації синтез РНК закінчується. Ще не ясно, як саме виявляється ця ділянка ДНК. Вважають, що вона містить особливу термінальну послідовність нуклеотидів, яка розпізнається термінуючим білком – ?-фактором. Після термінації фермент відокремлюється від матриці і може зв’язатись з новою молекулою ?-фактора і розпочати транскрипцію нової молекули РНК.
На одному опероні можуть одночасно працювати десятки молекул РНК-полімерази, які знаходяться в постійному русі, одна за одною, утворюючи велику кількість РНК-копій. Це доведено за допомогою електронних мікрофотографій ділянок генома в стані активної транскрипції.Утворені молекули РНК підлягають процесінгу. Це може бути розщеплення великої молекули-попередника на коротші. З попередників м-РНК вирізаються специфічно беззмістовні ділянки – інтрони. Ті ділянки РНК, які залишаються у складі дозрілої молекули, називаються екзонами. Відбувається ферментативна модифікація деяких груп, зокрема метилювання, приєднання кінцевих груп. Наприклад, до більшості м-РНК еукаріот на 5/-кінці приєднується “кеп” (з англ. “шапка”) - залишок 7-метигуанозину, приєднаний через незвичайний трифосфатний зв’язок Вважають, що “кеп”, як і інші пристосування, необхідний для захисту молекули від руйнування.
Зворотна транскрипціяПісля встановлення функції ДНК шлях спадкової інформації від ДНК до РНК, а від неї до білку був довгий час незаперечною догмою молекулярної біології. Тому відкриття в 1970 p зворотного шляху, від РНК до ДНК, і ферменту, який каталізує цей процес - РНК-залежної-ДНК-полімерази, довгий час сприймалось як абсурдне. Тим не менш, на наш час уявлення про існування процесу зворотної транскрипції, тобто синтезу ДНК на РНК-матриці, є цілком визнаним, доведеним і плідно використовується.
Зворотна транскрипція здійснюється при трансформації спадкової інформації від РНК РНК-ових (або ретро-вірусів) до ДНК в клітині-господарі. Новоутворена ДНК, комплементарна вірусній РНК, служить інформацією в клітині для синтезу нових вірусів. Вона реплікується тими ж ферментами, що і власна ДНК господаря. В незалежних дослідженнях Д. Балтімора і Т. Теміна в І969-І970 pp. були виділені ферменти РНК-залежні-ДНК-полімерази (ревертази або зворотні транскриптази).Процес зворотної транскрипції відбувається, зокрема, при зараженні онкогенними вірусами, вірусом СНІДу. При проникненні в еукаріетичну клітину, РНК-овий вірус забезпечує синтез ферменту - ревертази. Ревертаза каталізує зворотної транскрипцію, в результаті якої синтезується ланцюг ДНК. На ньому добудовується комплементарний ланцюг. Утворена дволанцюгова ДНК вбудовується в ДНК клітини господаря, або утворює самостійну молекулу в цитоплазмі (плазміду). Зчитування її інформації в кінцевому рахунку приводить до переродження клітини.
Таким чином, завдяки процесу зворотної транскрипції долається міжвидовий бар’єр в перенесенні спадкової інформації.Виявлення і дослідження механізму зворотної транскрипції відкриває великі теоретичні і практичні перспективи. Наприклад, визначення наявності ревертаз в організмі використовують при діагностиці деяких форм раку (при лейкемії). Висловлюються припущення про можливість створення універсальних антисивороток для пригнічення процесу оберненої транскрипції, а отже, і ракового переродження клітин.Ревертази стали важливим інструментом в молодій галузі біології - генній інженерії. Генну інженерію визначають як систему експериментальних прийомів, які дозволяють створювати в лабораторії, в пробірці штучні (рекомбінантні) генетичні структури. Завдяки оберненій транскрипції стало реальністю створення химерних генотипів, які поєднують гени різних видів. Вже здійснені роботи по створенню бактерій, які містять множинні гени синтезу цінних білків інтерферону, або гормону росту. Такі бактерії напрацьовують білок у великих кількостях, забезпечуючи ним практичні потреби.
Технологія таких синтезів відшліфовується. Відомості про створення все нових штучних поєднань генетичного матеріалу з’являються постійно. Наприклад, здійснено об’єднання гену фіксації азоту з хромосомою рослинної клітини. Виведення культур з таким генотипом дозволило б вирішити проблему забезпечення рослин азотними добривами.В перспективі генні інженери сподіваються виліковувати спадкові захворювання, замінюючи ушкоджений ген на здоровий. Однак виведення химерних генотипів може викликати непрогнозовані, катастрофічні для людства наслідки. Крім того існують проблеми етичного плану, адже генна інженерія здійснює втручання в тонкі механізми спадкової функції. І не виключено, що за такі експерименти візьмуться з винищувальною метою. Тому в різних країнах законодавче обмежуються деякі експерименти в галузі генної інженерії.
Список використаних джерел: Кучеренко М.Г. Загальна біологія 10 клас К: « Генеза»,2001р. Мердух І.І. Ладун Я.Д. Біологія . Довідник школяра.Тернопіль, 2001 р. Барна І.В. Загальна біологія. Пробний підручник.10 клас Тернопіль, 2009 р. Хімія: Посібник для вступників до вищих навчальних закладів/ В.В. Сухан, Т.В. А.Й. Капустян, В.Ф. Горлач. – 2-е вид. – К.: Либідь, 1995. – 448. www.Cell image library , www.youtube.com , www.uk.wikipedia.org
Виконала: Осадча оксана Миколаївна Вчитель біології та хімії Бойківщинської ЗОШ І-ІІІ ступенів