Download

1 / 1
10 likes | 162 Vues
Vestibular hair cell regeneration induced by math1 gene therapy. Hinrich Staecker3, Mark Praetorius1, Kim Baker3, and Doug Brough2 1Dept. of Otolaryngology, University of Saarland, Homburg Germany 2Genvec Inc., Gaithersburg MD

E N D
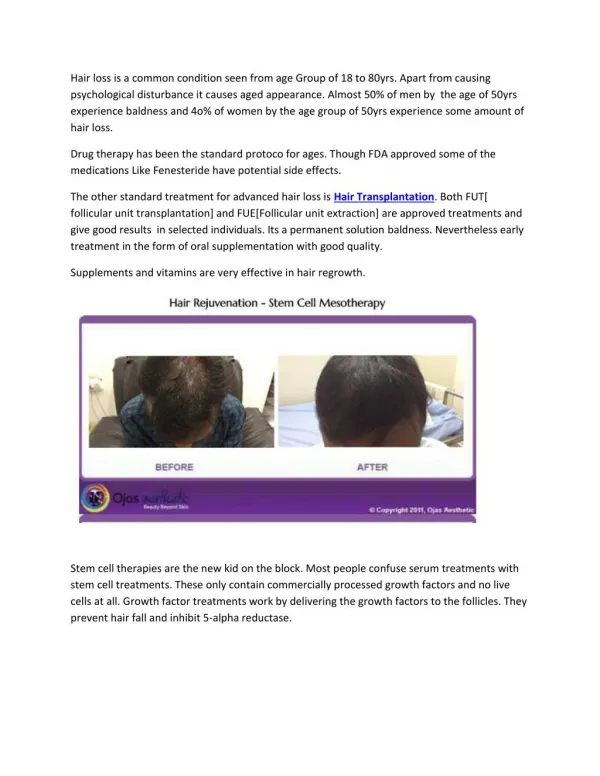
Vestibular hair cell regeneration induced by math1 gene therapy Hinrich Staecker3, Mark Praetorius1, Kim Baker3, and Doug Brough2 1Dept. of Otolaryngology, University of Saarland, Homburg Germany 2Genvec Inc., Gaithersburg MD 3Otolaryngology Head and Neck Surgery University of Maryland School of Medicine,, Baltimore, MD Figure 4 Abstract Over expression of math1 has been shown to induce generation of hair cells in the cochlea in neonatal tissue culture models of the organ of Corti as well as in the cochlea in vivo. We have examined an in vitro organotypic culture model as well as unilateral acute models of aminoglycoside vestibular injury. Delivery of math1 using an adenovector (Ad) resulted in the generation of hair cells in adult mouse utriclular and saccular cultures. Mice treated acutely with aminoglycosides showed recovery of a significant portion of the vestibular neuroepithelium after Ad math1 was infused into the inner ear. Assessment of animals 2 months after vector infusion demonstrated a recovery of swim times compared to non vector treated controls, demonstrating recovery of vestibular function after math1 gene therapy. Swim test results : Animals were swim tested one month after aminoglycoside ablation of one followed by no treatment or treatment with Ad math1. Age matched control animals showed a baseline swim test time of 10 + 1.4 s (Fig 4 ). Aminoglycoside only treated animals showed average swim test times of 22 + 4.2 s. Animals treated with aminoglycoside followed by Ad Math1 showed swim times of 12 + 3 s (p<0.01). Control animals showed an average hearing threshold of 40 dB whereas all animals treated with intracochlear injection of aminoglycoside showed hearing losses of >90 dB. Materials and Methods Figure 2 Results of Ad math 1 in animals undergoing unilateral aminoglycoside injury: Infusion of 2 μl of neomycin 10-3 M into the round window membrane reliably resulted in unilateral deafness and loss of all auditory and vestibular hair cells (Fig 2 C, D). Animals treated with Ad math1 after aminoglycoside treatment showed restoration of hair cells in the macular organs and the crista ampularis (Fig 2 A, B). Macular organ culture: Adult C57Bl6xCBA mice were anesthetized with intraperitoneal avertin and decapitatied. The otic capsule was exposed and the macular organs identified by finding the otolithic membranes. Using a #5 watchmaker forceps the saccule and utricle were removed and the otolith layer dissected away. The organs were cultured on a millicel membrane suspended in 500 μl of DMEM supplemented with N1+100 u/ml penecillin and glucose. After 24 hours in vitro (37 oC, 5% CO2) cultures were treated with neomycin 10-3 M for 48 hours. The cultures were washed in medium and then maintained for 4 weeks in vitro. Control cultures were left in medium that was exchanged every 7 days. Experimental cultures were treated with 1E9 particle units (pu) of E4 deleted adenovector carrying the gene for math1 driven by a CMV promoter (Ad11dmath1). Experimental cultures were similarly maintained in vitro for 4 weeks. At four weeks in vitro, explants were washed in phosphate buffered saline (PBS) and fixed in 4% paraformaldehyde. Explants were treated with PBS + 0.3% triton x-100 (Sigma) for 5 min and again washed in PBS followed by treatment with phalloidin-TRITC. Explants were washed in PBS x3 and mounted in antifade medium. Hair cell counts were achieved by placing a grid over the images that allowed counts of total stereocilia bundles per 0.1 mm2 area to be made in 5 separate regions of the explants. Animal surgery and vector treatment: Adult C57Bl6 mice were anesthetized using Avertin (IP). After anesthetizing the mouse the skin over the incision is shaved and sterilized with betadine solution. A dorsal postauricular incision is made and the bone medial to the tympanic ring exposed. Using an 18# needle a hole is drilled exposing the middle ear space medial to the tympanic ring. The round window niche is identified and the bone overhanging the niche identified. The bone is gently scraped away revealing the round window membrane. The animals are then mounted in a stereotactic headframe using a snout holder. Animals undergoing unilateral lesioning are injected with 2 μl of neomycin 10 -3 M. The cochleostomy is then sealed and the animals allowed to recover for 48 hours. Vector injections consisted of 1E8 PU of Ad11dmath1 (n=10). Vector injections were carried out using a Hamilton microsyringe with 0.1 µl graduations. A custom made 36 gauge needle will be used to puncture the round window membrane. The injection was carried out using a micromanipulator (Singer Instruments, England) while the animals were immobilized to minimize injection trauma. After the injection the round window is patched with a small piece of muscle. A matched number of control animals were injected with 1 µl of artificial perilymph. Hearing and Balance testing: ABRs are performed using a Tucker Davis System 3. Mice are anesthetized and maintained on a heating pad. Needle electrodes will be placed on the ipsilateral mastoid tip and at the vertex. Precalibrated stimuli ranging from 500 to 24, 000 Hz are presented from 90 to 10 dB in 5 dB steps. An average of 1000 stimuli are collected. Animals were tested for a change in balance function by swim testing. Animals were dropped into a opaque high sided pool of body temperature water from a hight of 20 cm. Time in seconds between the animals contact with the water and purposeful swimming was noted. Histology: Animals were anesthetized with intraperitoneal avertin and sacrificed via intracardiac perfusion with 4% paraformaldehyde. The temporal bones are then removed and the stapes removed. The temporal bones are postfixed in 4% paraformaldehyde in PBS overnight at 4oC. After rinsing in PBS three times for 30 minutes, the temporal bones were decalcified in in Calex (Fisher) for 48 hrs. After rinsing again the temporal bones were dehydrated, cleared and embedded in paraffin. Seven µm serial sections are mounted on sialanated slides (Sigma). The sections are then deparaffinized and rehydrated. Sections will be stained by standard hematoxilin and eosin methods, dehydrated and mounted. Images are captured with a digital camera (Spot Technologies). Total hair cells for the utricle and saccule were determined by counting every 5th section. Introduction Replacement or repair of vestibular hair cells has been demonstrated in a variety of model systems including guinea pig macular organs (Forge et al. 1616-19), rodent and human macular organs (Warchol et al. 1619-22), and the chinchilla crista ampullaris (Tanyeri, Lopez, and Honrubia 194-202). A variety of factors were identified that could increase the number of hair cells generated in an aminoglycoside injury/regeneration-repair model including retinoic acid, TGF α and IGF (Kelley, Talreja, and Corwin 3013-26;Lambert 701-18;Oesterle and Rubel 34-46;Kopke et al. 5886-91). Kopke et. al. demonstrated that infusion of a cocktail of growth factors after aminoglycoside injury in guinea pigs resulted in a statistically significant renewal of vestibular hair cells and recovery of vestibular function as measured by recovery of the horizontal vestibule-occular reflex (VOR)(Kopke et al. 5886-91). Particularly significant was the demonstration that VOR phase recovered indicating that recovery of balance function in this animal was not due to central compensation. A variety of studies have suggested that the proliferation observed in many of these models may represent differentiation of supporting cells or even repair of hair cells rather than replacement of hair cells from a mitotic event (Li and Forge 433-46). The effort to determine the molecular basis for this observed effect has led to examination of factors controlling hair cell differentiation (Zine et al. 4712-20;Shailam et al. 809-19). One of the genes that appears to be important for the generation of hair cell is the mammalian atonal homolog math1. Mice carrying a homozygous knockout of math1 fail to develop auditory or vestibular hair cells (Bermingham et al. 1837-41). Delivery of a plasmid vector expressing math1 has been shown to produce supernumerary hair cells in vitro (Zheng and Gao 580-86). These results were repeated using the human homolog of math1 (hath1) delivered via an adenovector (Shou, Zheng, and Gao 169-79). Most recently, generation of new auditory hair cells in the mouse cochlea was demonstrated using math1 delivered by an adenovector (Kawamoto et al. 4395-400). Since the vestibular epithelium has a strong potential for spontaneous regeneration we wanted to determine the feasibility of restoring vestibular hair cells and vestibular function in a mouse model of unilateral aminoglycoside ototoxicity. Figure 1 Discussion Fluorescent photomicrographs of macular explants treated with neomycin and Ad11D math 1. Panel A demonstrates lack of regeneration of stereocilia in explants left in vitro after aminoglycoside injury. Explants 1 week after Ad Math1 are shown in B. At this point there are no clearly identifiable stereocilia. After 4 weeks post math 1 gene therapy randomly oriented stereocilia can be seen (C). A control explant can be seen in D. Control treated cultures showed an average of 39 + 4 stereocilia bundles per 0.1 mm2. Aminoglycoside only treated explants showed 6 + 3 stereocilia bundles per 0.1 mm2. Explants treated with aminoglycoside followed by Ad math1 demonstrated presence of sterociliary bundles at 29 + 7 bundles per 0.1 mm2. The Ad math1 treated cultures showed a statistically significant counts of stereocilia compared to aminoglycoside only treated cultures (p<0.01). A number of studies have demonstrated that the vertebrate vestibular epithelium has the potential to regenerate after loss of hair cells induced by aminoglycoside injury (Forge et al. 1616-19;Lambert 701-18;Tanyeri, Lopez, and Honrubia 194-202). These experiments demonstrated that the vertebrate inner ear had the potential to spontaneously recover from an injury or recover with the aid of a variety of growth factors that induced cell differentiation and proliferation. Kopke et. al. demonstrated that a cocktail of growth factors infused into the ear of an ototoxin treated guinea pig via an Alzet™ osmotic pump resulted in significant recovery of stereociliary bundles and a recovery of vestibular function measured via the horizontal VOR (Kopke et al. 5886-91). Discovery of math 1, the gene that controls development of auditory and vestibular hair (Bermingham et al. 1837-41)cells has renewed attempts to induce the regeneration of hair cells in the mammalian inner ears. Application of a gene transfer approach has the advantage of needing only a single application of vector to achieve renewal of hair cells. Transfer of a plasmid vector expressing math1 generated new hair cells in neonatal rat inner ear cultures (Zheng and Gao 580-86). This effect was also demonstrated using the human homolog of math1, hath1 when applied to adult rat maculae using an adenoviral vector. (Shou, Zheng, and Gao 169-79) . Interestingly Brdu labeling suggested that the hair cells generated after hath1 gene therapy were not the result of a cell division. Most recently it was demonstrated that infusion of Ad math1 into the scala media resulted in regeneration of auditory hair cells (Kawamoto et al. 4395-400). In this series of experiments we have demonstrated that use of an adenovector can be used to effectively deliver math1 to the adult mammalian vestibular system in vitro or in vivo. Macular cultures showed recovered approximately 80% of their stereocilia after one month in vitro consistent with previously described studies (Shou, Zheng, and Gao 169-79). In vivo studies in which a 2 μl volume of vector was delivered showed recovery of vestibular function, as determined by swim testing and histologically recovered 80% of the damaged vestibular neuroepithelium. In our study we did not note any recovery of hair cells in the auditory system even though prior studies suggest that an injection of adenovector should be distributed throughout the inner ear (Praetorius et al. 211-14;Staecker et al. 157-63). Kawamoto et. al. demonstrated that injection of the vector into the endolympatic space was required to demonstrate regeneration of hair cells in the cochlea (Kawamoto et al. 4395-400). The biologic basis of this observed phenomenon is not clear but may relate to differences in lateral inhibition in the vestibular and auditory system or possibly due to differences in the animal models used. The vestibular system is a particularly important target for hair cell regeneration since currently no clinical treatments are available for patients that have lost all vestibular function. Complete hearing loss can at present be treated with amplification or cochlear implantation depending on the severity of the loss . At present there is no vestibular equivalent to a cochlear implant and the engineering challenges to creating one are considerable. Several studies have now demonstrated the effectiveness of hair cell regeneration using gene transfer of the math1 gene. The vestibular system as a whole may have a latent potential for repair, explaining the occasional report of recovery of balance function in aminoglycoside ototoxicity in humans. However this series of experiments demonstrates that addition of math1 after aminoglycoside injury significantly enhances hair cell recovery. Follow up experiments will define the VOR response in mice undergoing vestibular hair cell regeneration and define the time lag between injury and potential regeneration. Sponsored by a research grant from Genvec Inc. and the Maryland Industrial Partnership foundation Figure 3 Counts of serially sectioned macular organs showed average hair cell counts of 11.8 + 2 for untreated controls (n=5), 1.2 + 2 (n=10) for aminoglycoside only treated animals and 8.1 + 2.2 for aminoglycoside and Ad math1 treated animals (Fig 3). This represents a statistically significant recovery of hair cells in animals treated with aminoglycoside and Ad math1 compared to aminoglycoside only treated animals (p<0.01). Both type I and type II hair cells were noted in the Ad math1 treated vestibular epithelium. References • Bermingham, N. A. et al. "Math1: an essential gene for the generation of inner ear hair cells." Science 284.5421 (1999): 1837-41. • Forge, A. et al. "Ultrastructural evidence for hair cell regeneration in the mammalian inner ear." Science 259.5101 (1993): 1616-19. • Kawamoto, K. et al. "Math1 gene transfer generates new cochlear hair cells in mature guinea pigs in vivo." J.Neurosci. 23.11 (2003): 4395-400. • Kelley, M. W., D. R. Talreja, and J. T. Corwin. "Replacement of hair cells after laser microbeam irradiation in cultured organs of corti from embryonic and neonatal mice." J.Neurosci. 15.4 (1995): 3013-26. • Kopke, R. D. et al. "Growth factor treatment enhances vestibular hair cell renewal and results in improved vestibular function." Proc.Natl.Acad.Sci.U.S.A 98.10 (2001): 5886-91. • Lambert, P. R. "Inner ear hair cell regeneration in a mammal: identification of a triggering factor." Laryngoscope 104.6 Pt 1 (1994): 701-18. • Li, L. and A. Forge. "Morphological evidence for supporting cell to hair cell conversion in the mammalian utricular macula." Int.J.Dev.Neurosci. 15.4-5 (1997): 433-46. • Oesterle, E. C. and E. W. Rubel. "Hair cell generation in vestibular sensory receptor epithelia." Ann.N.Y.Acad.Sci. 781 (1996): 34-46. • Praetorius, M. et al. "Hearing preservation after inner ear gene therapy: the effect of vector and surgical approach." ORL J.Otorhinolaryngol.Relat Spec. 65.4 (2003): 211-14. • Shailam, R. et al. "Expression of proneural and neurogenic genes in the embryonic mammalian vestibular system." J.Neurocytol. 28.10-11 (1999): 809-19. • Shou, J., J. L. Zheng, and W. Q. Gao. "Robust generation of new hair cells in the mature mammalian inner ear by adenoviral expression of Hath1." Mol.Cell Neurosci. 23.2 (2003): 169-79. • Staecker, H. et al. "Gene expression in the mammalian cochlea: a study of multiple vector systems." Acta Otolaryngol. 121.2 (2001): 157-63. • Tanyeri, H., I. Lopez, and V. Honrubia. "Histological evidence for hair cell regeneration after ototoxic cell destruction with local application of gentamicin in the chinchilla crista ampullaris." Hear.Res. 89.1-2 (1995): 194-202. • Warchol, M. E. et al. "Regenerative proliferation in inner ear sensory epithelia from adult guinea pigs and humans." Science 259.5101 (1993): 1619-22. • Zheng, J. L. and W. Q. Gao. "Overexpression of Math1 induces robust production of extra hair cells in postnatal rat inner ears." Nat.Neurosci. 3.6 (2000): 580-86. • Zine, A. et al. "Hes1 and Hes5 activities are required for the normal development of the hair cells in the mammalian inner ear." J.Neurosci. 21.13 (2001): 4712-20.