Download

1 / 45
450 likes | 620 Vues
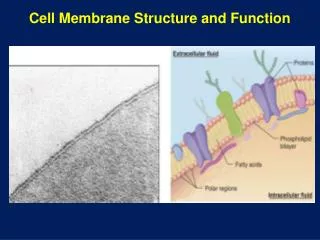
Membrane Transport “Pores, Porters and Pumps”. CH353 March 4-6, 2008. Summary. Thermodynamics and Kinetics of Membrane Transport Classification of Membrane Transport Proteins Channels, Porters, Primary Active Transporters Primary Active Transporters

E N D
Membrane Transport“Pores, Porters and Pumps” CH353 March 4-6, 2008
Summary • Thermodynamics and Kinetics of Membrane Transport • Classification of Membrane Transport Proteins • Channels, Porters, Primary Active Transporters • Primary Active Transporters • driven by hydrolysis of phosphoanhydride bonds • Porters (secondary active transport & facilitated diffusion) • driven by electrochemical potential • Systems combining active transporters and porters • Channels (for water and ions) • Regulation of ion channels • voltage and ligand gating • action potential and synaptic function
Non-MFS Porters • Na+ Ca2+ Exchanger (NCX1) • member of Ca2+: cation antiporter (CaCA) family (2.A.19) • antiporter of 3 Na+ for 1 Ca2+ • Na + Glucose Symporter • member of solute: sodium symporter (SSS) family (2.A.21) • symporters of 1-2 Na+ for 1 glucose • HCO3– Cl– Exchangers • members of anion exchanger (AE) family (2.A.31) • antiporters and symporters
Na+ Ca2+ Exchanger (NCX1) • Plasma membrane antiporter regulating intracellular Ca2+ in cardiac myocytes • Electrogenic process: transports 3 Na+ in for 1 Ca2+ out • Lower affinity for Ca2+ than Ca2+ ATPase pumps, but faster • 2000 Ca2+ /sec vs. 30 Ca2+ /sec • Lowering Ca2+ reduces strength of heart muscle contraction • Digoxin inhibits Na+K+ ATPase increasing intracellular Na+, decreasing efficiency of Na+ Ca2+ exchanger, increasing strength of heart contraction Na+ Ca2+ Exchanger regulated by cytoplasmic Ca2+ binding domain CBD1 on exchanger Blaustein et al. 2007, Proc Natl Acad Sci USA 104: 18349
Na+ Glucose Symporters • secondary active transport of glucose up concentration gradient coupled with Na+ down electrochemical gradient (electrogenic) • 2 human isoforms: • SGLT1 (SLC5A1): high affinity, low capacity • symport of 2 Na+ in for 1 glucose in • intestinal and renal absorption of glucose and galactose • SLC5A1 deficiency: glucose-galactose malabsorption (GGM) • SGLT2 (SLC5A2): low affinity, high capacity • symport of 1 Na+ in for 1 glucose in • renal glucose reabsorption • SLC5A2 deficiency: renal glucosuria
Glucose Transport in Intestinal Epithelium • Apical: SGLT1 • Basolateral: GLUT2 and Na+K+ ATPase Molecules absorbed from intestinal lumen pass through epithelial cells Transporters in epithelial cells require correct localization on apical or basolateral membranes Tight junction
C2 C1 ∆G (Na+ transport) = RT ln()+ ZF∆y = -6.4 kJ/mol + -4.9 kJ/mol = -11.3 kJ/mol For 1 mol Na+ transported per mol glucose: 11.3 kJ/mol = RT ln ( ) [glucose]in [glucose]out = 80.7 [glucose]in [glucose]out [glucose]in [glucose]out For 2 mol Na+ transported per mol glucose: 22.6 kJ/mol = RT ln ( ) [glucose]in [glucose]out = 6520 Energetics of Na+ Glucose Transport • Secondary active transport of glucose coupled with the electrochemical gradient of Na+ is energetically favorable Na+out + glucoseout→ Na+in + glucosein [Na+]out = 145 mM [Na+]in = 12 mM ∆y = - 50 mV At equilibrium: ∆G = ∆G (Na+ transport) + ∆G (glucose transport) = 0
Anion Exchangers HCO3– Cl– Exchangers (Antiporters) • 3 human members AE1 (SLC4A1) (erythrocytes) • binds to carbonic anhydrase (transport metabolon) • exchange of CO2 at tissues and lungs AE2 (SLC4A2) • intracellular pH regulation; linked to Na+ H+ exchanger • exocrine secretion AE3 (SLC4A3) • isoforms in kidney and heart
Regulation of Intracellular pH • Respiring cell accumulate excess acids, e.g. H2CO3 • 3 transport systems combine to regulate pH • Na+ H+ antiporter (↑ pH) • Na+ in; H+ out • Na+ HCO3– Cl– (↑ pH) • Na+, HCO3– in; Cl– out • HCO3– Cl– antiporter (↓ pH) • HCO3– out; Cl– in Intracellular pH activates appropriate transport system
( D-valine – L-lactate – L-valine – D-hydroxyisovalerate ) 3 Ionophores Non-ribosomally synthesized porters • Cyclic compounds that collapse membrane concentration gradients • Diverse composition (peptides, polyketides, polyethers, tetrolides) • Valinomycin – cyclic peptide K+ uniporter (K+ > Cs+ > Na+) • Monesin – cyclic polyketide Na+:H+ antiporter (Na+ > K+> Rb+) • Nigericin – cylic polyketide K+:H+ antiporter (K+ > Rb+ > Na+) • Tetronasin – cylic polyketide Ca2+:H+ or Mg2+:H+ antiporter Valinomycin – K+ complex
Experimental Methods for Transporters Models • Intact cells • Liposomes with purified or recombinant proteins • Oocyte expression of injected RNA Measurements • Membrane potential • Ion current (patch clamp)
Study Models for Transport Proteins • Isolate cell membrane fraction • Purify transporters from other integral membrane proteins • Mix solubilized transporter prep with phospholipids • Remove detergent and form liposomes with transporters • Use transporters in liposome for experiments
Study Models for Transporters • Harvest oocytes from Xenopus laevis (oocytes are ~1 mm dia) • Transcribe in vitro RNA from a cDNA construct • Microinject RNA into Xenopus oocytes • Allow for expression of RNA into protein • Measure properties of transporter [online]
Measurement of Transporter Activity Membrane potential is measured using: • microelectrode inserted into cell, • reference electrode in medium • potentiometer for measuring voltage and direction of current • System is used for measuring action potentials on neurons and muscle cells
Measurement of Transporter Activity Patch clamping technique • micropipette filled with electrolyte is placed over transporter on membrane • reference electrode in medium or inserted in cell • measure current required for maintaining constant a membrane potential • applied for measuring transport of ions
Measurement of Transporter Activity • Variations on experimental setup for patch clamping muscle cell or neuron
Measurement of Transporter Activity Interpreting patch clamping data • inside-out patches of muscle cell membrane clamped at voltage slightly less than resting potential. Drops in current (pA) indicate opening of single Na+ channels • patches of neuron membrane clamped at 3 different voltages. Increases in current show opening of K+ channels. Channel opening increases with membrane depolarization
Channels/Pores (α-Type Channels) Characteristics • Rapid transport without carriers near diffusion limit (107 to 109 /s) • Selectivity: molecule, charge or ion specific • Gating: open, closed, or inactivated
Aquaporins • Water-specific membrane channels • Members of major intrinsic protein (MIP) family (1.A.8) • Aquaporin group permeated by water only • AQP0, AQP1, AQP2, AQP4, AQP5, (AQP6 and AQP8) • Aquaglyceroporin group permeated by water and small solutes, e.g. anions, glycerol, urea • AQP3, AQP7, AQP9 and AQP10 • Different permeabilities to water • AQP0, AQP6, AQP9 and AQP10 have low permeability to water • Tissue specific expression of aquaporins: types, proteins /cell and cellular localization
Regulation of Aquaporins In kidney • AQP1 expressed in epithelial cells of proximal tubules; absorbs water from glomular filtrate • AQP2 expressed in principal cells; absorbs water from collecting ducts • Vasopressin, a antidiruetic hormone, stimulates urine concentration by translocating AQP2 from intracellular vesicles to the apical plasma membrane of principal cells
Structure of Aquaporins • AQP1 is a tetramer of 28-kDa subunits, each with a transmembrane channel 0.2 – 0.3 nm in diameter • each subunit has 6 transmembrane helices and 2 short helices (non-spanning) • each short helix extends from opposite sides toward the middle and has an Asn-Pro-Ala (NPA) motif • NPA motifs and connecting loops form a selectivity filter • the 2-nm x 0.28 nm diameter pore is selective for water and allows sequential passage of several waters; H-bonds formed with carbonyl oxygens; closely-spaced waters not permitted • hydrogen ions (H3O+) cannot pass through water channel; repelled by Arg and His residues and helical dipoles
Aquaporin Selectivity Filter • Structure of AQP1 subunit with amino acids lining water channel • 4 water molecules are indicated by green spheres • Constriction region (blue arrow) • Pseudo two-fold symmetry axis between N78 and N194 (black arrow) • Amino acids H76, H182, R197 prevent H3O+ entry • 4 water molecules too far apart for “H+ hopping” Sui et al. 2001, Nature 414: 872
Ion Channels Regulation of ion channels • Ligand gating • Voltage gating (some are refractory after opening) Ashcroft 2006, Nature 440: 440
K+ Channels Non-gated K+ channel (KcsA) (Streptomyces lividans) • Tetramer of same subunits each with • 2 transmembrane helices • a short helix • an ion-sensing loop (selectivity filter) (yellow) • Loops from each subunit (4) combine to form binding sites for 4 unsolvated K+ (green)
K+ Channel Selectivity Filter Extracellular Side • Carbonyl oxygens coordinate 2 unsolvated K+ at one time; each K+ is coordinated by 8 oxygens • K+ can occupy • positions 1 and 3 (blue) or • positions 2 and 4 (green) • Binding of Na+ (radius 0.95 Å) is energetically less favorable than binding of K+ (1.33 Å) • Transport of K+ is 10,000 times faster than that of Na+ 4 3 2 1 Intracellular Side
4 3 2 1 K+ Channel Transport Mechanism • Process starts with 2 K+ bound to the selectivity filter (sites 2 & 4) • K+ enters channel from cytosol • K+ loses hydration near the selectivity filter; is stabilized by dipoles of short α helices • K+ enters selectivity filter (site 1) displacing the K+ at site 2 to site 3 and the K+ at site 4 to outside; extracellular K+ is hydrated • K+ ions at sites 1 & 3 shift to sites 2 & 4 for next K+ to enter
Electrochemical Membrane Potential • Na+K+ ATPase establishes chemical gradients across the plasma membrane with high [K+] and low [Na+] inside • Secondary active transport using the Na+ gradient creates the gradients of other ions • Ca2+ also has active transport • Electrogenic transport of 3 Na+ out for 2 K+ in causes some of the membrane potential • Most of the membrane potential results from transport of K+ out of the cell through K+ channels
Resting Membrane Potential • Membrane potential is caused by the predominant flow of K+ from the cell • Most animal cells maintain their membrane potential at -50 to -70 mV relative to outside the cell • Excitable cells (neurons and muscle cells) transiently alter their membrane potential, making it more positive • Opening Na+ channels makes the membrane potential more positive (depolarizing the membrane) • Opening K+ or Cl– channels makes the membrane potential more negative (hyperpolarizing the membrane)
Action Potentials • Membrane potential of excitable cells becomes transiently more positive (depolarizes) in response to a stimulus – action potential • Action potential coincides with a brief increases in permeabilities of membrane to Na+ and K+ • Flow of Na+ out of cell depolarizes membrane; subsequent flow of K+ out of cell repolarizes and slightly hyperpolarizes membrane • Action potentials are caused by sequential opening and closing of voltage-gated Na+ and K+ channels
Transmission of Action Potential • Local depolarization of membrane spreads to adjacent region (purple) opening voltage-gated Na+ channels • This causes further depolarization of membrane and opening of additional voltage-gated Na+ channels • Na+ channels in the region of the membrane (green) where the action potential originated remain closed • These channels are refractory to opening by depolarization for several milliseconds • This results in the unidirectional transmission of the action potential
Model for Voltage-Gated Na+ Channel Experimental Support: • voltage-gated Na+ channel is bell-shaped by electron microscopy • opening of channel coincides with measured displacement of charged amino acids (gating charge) – movement of positively charged S4 helices • mutation of inactivating sequence or its binding site abolishes refractory period
S4 Family of Pore Loop Channels • Voltage-gated ion channels (S4 family) are related to the non-gated K+ channel (homologous S5 – pore loop – S6) • Voltage-gated K+ channels are tetramers of same 4 subunits • Voltage-gated Na+ and Ca2+ channels have 4 domains on one subunit
Topology of Voltage-Gated Na+ Channel • Large subunit (~260 kDa) has 4 homologous domains each with • 6 transmembrane helices (predicted by hydrophobicity) • a selectivity filter (pore loop), a voltage sensor (S4) and an activation helix (S6) • Inactivation sequence is between domains III and IV
Voltage Gating of Na+ Channels • Nice Model, But … No 3D Structural Support
Models for Movement of Gating Charges • Gating charges (red +) move from inside (bottom) to outside (top) with depolarization of membrane Conventional Model • Gating charges pass through the protein by translation or rotation of S4 helix within a gating pore of the protein New Model • Gating charges pass through the membrane by movement of voltage-sensor paddles on the outside of the protein Closed Open Jiang et al. 2003, Nature 423: 33
Structure of Voltage-Gated K+ Channel • 3D structure of voltage-gated K+ channel shows voltage sensor on a paddle – a helix-loop-helix formed by S3b and S4 • gating charges are shown as blue side chains • paddle is outward in open form and rotates inward in closed form (displacement of ~15 Å) • this pushes down on the S4-S5 linking helix (orange) causing it to tilt down • this presses on the S6 helix (blue) closing the channel Long et al. 2007, Nature 450: 376
Ligand Gated Ion Channels • Cys-loop family of receptors • Pentmeric receptor complexes with a central channel • 5 subunits are homologous to each other • Cationic channel receptors • Allow entry of Na+ and Ca2+ (negative charges in channel) • Depolarizes membrane (excitory) • Acetylcholine and Serotonin are ligands for receptors • Anionic channel receptors • Allow entry of Cl– and HCO3– (positive charges in channel) • Hyperpolarizes membrane (inhibitory) • γ-aminobutyrate (GABA) and glycine are ligands for receptors
Nicotinic Acetylcholine Receptor • Model of acetylcholine receptor (Torpedo electric organ) • Based on EM data on receptor and X-ray structure of binding protein • Pentameric receptor a2bgd with each subunit going the length of receptor • each subunit has 3 domains • Extracellular β-sheet domain with 2 acetylcholine binding sites (on a2) • Channel domain of 4 transmembrane α-helices (M2 helix forms channel) • Cytoplasmic domain of α-helix a subunits are orange; ion channel is dark blue Sine & Engel 2006, Nature 440: 448
K1* K2* open R* AR* A2R* θ0 A θ1 A θ2 AR A2R closed R K1 K2 Ligand Binding and Channel Conformations K1*, K2* ~ 109 θ2 / θ0= 107 K1, K2 ~ 106 • Receptor alternates between closed and open conformations • Acetylcholine has high affinity (nanomolar Kd) for open receptors and low affinity (micromolar Kd) for closed receptors • Equilibrium constant between closed and open forms, θ, is 107 times higher for receptor with 2 ligands θ2 than for unligated receptor θ0
Acetylcholine Receptor Mechanism • Acetylcholine binding ~50 Å away causes 15º rotation of M2 helix • Channel widens from 3 Å (closed) to 8 Å (open) in diameter • Open pore allows passage of hydrated cation (~8 Å in diameter)
Ion Channels in Neural Transmission • Ion flow through non-gated K+ channels polarizes membrane (– inside) • Depolarization opens then inactivates voltage-gated Na+ channels (+ inside) • Depolarization opens then inactivates voltage-gated K+ channels (– inside) • Action potential propagation opens then inactivates more voltage-gated Na+ and K+ channels (– to + to – inside) • Depolarization at axon end opens voltage-gated Ca2+ channels ([Ca2+] to > 1 μM) • Ca2+ activates SNAREs causing fusion of acetylcholine vesicle with membrane; the neurotransmitter diffuses across synapse
Post-synaptic Neuron Acetylcholine binds to receptor opening ligand-gated cation channel which depolarizes membrane (+ inside) Depolarization opens voltage-gated Na+ and K+ channels, stimulating the post-synaptic neuron This may result in transmission of action potential in the post-synaptic neuron Muscle cell Acetylcholine binds to receptor opening ligand-gated cation channel, which depolarizes membrane (+ inside) Depolarization opens voltage-gated Ca2+ channels raising [Ca2+] to > 1 μm Elevated [Ca2+] activates actin and myosin interaction causing muscle contraction Depolarization opens voltage-gated Na+ and K+ channels, propagating action potential on muscle cell plasma membrane Ion Channels in Neural Transmission
Inhibitors of Ion Channels • Voltage-gated Na+ channels tetrodotoxin (puffer fish) saxitoxin (Gonyaulax) • Voltage-gated K+ channels dendrotoxin (black mamba snake) • Voltage-gated Ca2+ channels w-conotoxin (cone snail) • Acetylcholine receptors a-conotoxin tubocurarinecobratoxin bungarotoxin (Kd = 10-15)