Download
1 / 45
450 likes | 459 Vues
The plasma membrane is a thin, selective barrier that serves as a protective barrier for cells. It contains sensors, channels, and pumps, and has remarkable mechanical properties. This text explores the composition and function of the lipid bilayer, membrane fluidity, and asymmetry, as well as the synthesis and modification of membranes in eukaryotic cells.

E N D
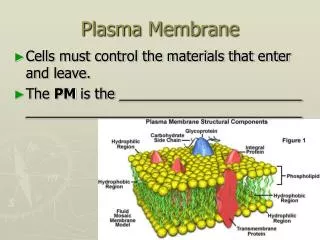
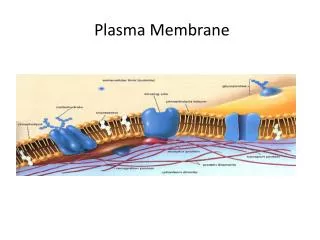
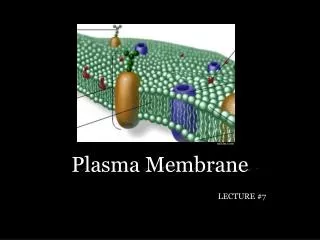
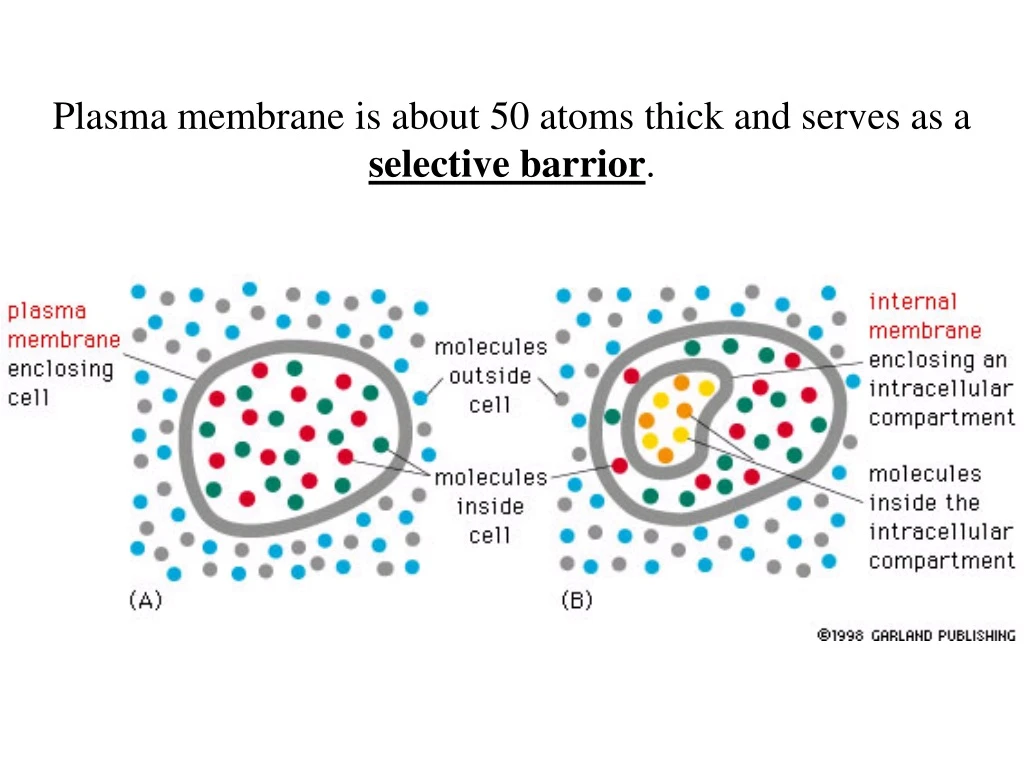
Plasma membrane is about 50 atoms thick and serves as a selective barrior.
Membranes include 1. sensors which enable the cell to respond to the environment and 2. highly selective channels and pumps. Mechanical properties of the membranes are remarkable. Enlarges and changes shape as needed with no loss of integrety.
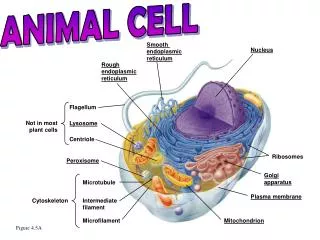
Eucaryotic cells contain many compartments created by intracellular membranes
The lipid bilayer. A. An electron micrograph
Phosphatidylcholine is the most common type of phospholipid. Positive negative
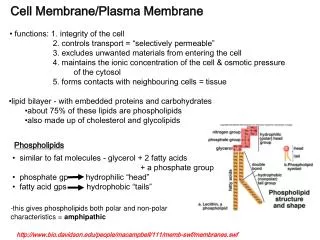
Three kinds of membrane lipids, all amphipathic, incude phospholipids, sterols, and glycolipids. Hydrophilic heads
Hydrophobic molecules in water. Water molecules form a more ordered structure. See question 1.
Purely hydrophobic molecules coalesce into a single drop in water. Amphipathic molecules like phosphotidylethanolamine form a lipid bilayer - energetically most favorable.
Self-sealing property “free edges” are quickly eliminated because they are energetically unfavorable - hydrophobic areas are in contact with water. The lipids will spontaneously seal and will always form a closed compartment. A small tear will be repaired. A larger tear may lead to the break up of the membrane into separate vesicles.
With water inside and out, the lipid bilayer remains intact, no lipids leave. However, the lipids do move freely within the bilayer. Experiments use liposomes, which form spontaneously.
Or flat bilayers formed across a hole in a partition between to aqueous compartments. The fluidity of the lipid bilayer is crucial for the function of the membrane. In these experimental systems phospholipids very rarely flip from one layer to the other without proteins to facilitate the process.
Due to thermal motions, lipid molecules within a monolayer rotate very rapidly and diffuse rapidly through the fluid membrane. Any drop in temperature decreases the rate of lipid movement, making the lipid bilayer less fluid. This inhibits many functions of the cell’s membranes. All this has been confirmed in whole cells.
The fluidity of a lipid bilayer depends on its composition. • As temperature and environment changes, the fluidity of the cell’s membranes must be kept functional. • The closer and more regular the packing of the tails, the more viscous and less fluid the bilayer will be • The length and degree of saturation with hydrogens affect their packing • shorter tails can not interact as much - more fluid • one of the two hydrocarbon tails often has a double bond - unsaturated. This creates a kink - less packing, more fluid.
Plant fats are generally unsaturated and liquid at room temperature. Animal fats are solid at room temperature. Hydrogenated plant fats are no longer unsaturated.
In bacterial and yeast cells, both the lengths and the unsaturation is constantly adjusted to maintain the membrane at a relatively constant fluidity. • At higher temperatures the cell makes longer tailed lipids with fewer double bonds. • In animal cells, membrane fluidity is modulated by cholesterol, which is absent in plants, yeast and bacteria.
Cholesterol fills in the spaces left by the kinks; stiffens the bilayer and makes it less fluid and less permeable.
Membrane fluidity is important to a cell for many reasons. • 1. Enables membrane proteins to diffuse rapidly and interact with one another - crucial in cell signaling etc. • 2. Provides a simple means of distributing membrane lipids and proteins by diffusion from sites of insertion. • 3. Allows membranes to fuse with one another and mix their molecules • 4. Ensures that membrane molecules are distributed evenly between daughter cells. • Remember though, cell has control - cytoskeleton and other interactions can limit the mobility of specific lipids and proteins.
The lipid bilayer is asymmetrical, with the cytoplasmic side being different from the non-cytoplasmic side. Proteins are embedded with a specific orientation crucial for their function. Phospholipid composition also varies.
New phospholipid molecules are synthesized in the ER by membrane-bound enzymes which use substrates (fatty acids) available only on one side of the bilayer. Flipases transfer specific phospholipid molecules selectively so that different types become concentrated in the two halves. One sided insertion and selective flippases create an asymmetrical membrane
In eucaryotic cells nearly all new membrane synthesis occurs in the ER. The new membrane is exported to the golgi apparatus for modification and export. Carbohydrate chains are added in the golgi - glycolipids. The enzymes that add sugar groups to lipids are confined to the golgi apparatus and sugars are added only to the non-cytoplasmic side. No flippases exist for glycolipids. Forms a protective coat on most animal cells. Intracellular signal transduction
Lipids are made in the ER and transported via vesicles to their destination. This form of transport preserves the cytoplasmic face and the non-cytoplasmic face which is exposed to the exterior of the cell or the interior of an organelle.
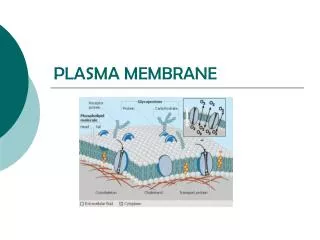
Lipid bilayers are impermeable to solutes and ions. Rate of diffusion varies depending on size and solubility properties. This has been demonstrated in synthetic bilayers. In this way, cells control the passage of molecules across its membranes Specialized transport proteins transfer specific substrates across the membrane
A crucial function of any cell membrane is to act as a barrior and to control the passage of molecules across it. The proteins in membranes serve many functions besides transporting nutrients etc. Linkers link intracellular actin filaments to extracellular matrix proteins. Receptors bind hormones and other signaling molecules and transmit that signal to the interior of the cell. Enzymes catalyze specific reactions - example: flippases.
Membrane proteins associate with the lipid bilayer in three main ways. 1. 2. 3. All membrane proteins have a unique orientation - a particular section always facing the cytosol. This is a consequence of how they are made. Integral membrane proteins (transmembrane and lipid-linked) can be isolated from the lipid membrane only by harsh treatment (detergent) whereas peripheral membrane proteins can be released by relatively gentle extraction methods.
Membrane proteins have a unique orientation. Always has the same region of the protein facing the cytosol. This depends on how it was made. Transmembrane proteins usually cross the bilayer as alpha-helices. Sometimes as beta-barrels
Transmembrane portions are composed largely of amino acids with hydrophobic side chains. However, the peptide back bone (peptide bonds) is hydrophilic. Therefore, a helical structure is the most energetically favorable.
The peptide bonds are hydrogen bonded to each other in the interior while the hydrophobic amino acid side chains contact the lipid chains.
Many transmembrane proteins cross the membrane only once. Many receptors for extracellular signals include an extracellular portion which binds the signal molecule (hormone etc.). Binding of the signal molecule induces a change in shape in the cytoplasmic part, which then signals to the cell’s interior.
Other transmembrane proteins form aqueous pores that allow water-soluble molecules to cross the membrane. These are more complicated, often cross the bilayer a number of times as alpha-helices or as beta-barrels In these cases alph-helices contain both hydrophobic and hydrophilic amino acid side chains, with the hydrophobic side chains on one side and hydrophilic on the other side.
Although the alpha-helix is the most common, transmembrane portions of a protein can be beta-barrels (two beta-sheets connected by a disulfide bond. The loop areas often form the active site or binding site. Beta-barrels are less versatile since the can form only wide channels.
To study proteins, they can be isolated from the lipid bilayer by solubilization using detergents. Detergents are small, amphipathic, lipidlike molecules with only a single hydrophobic tail. The protein-detergent complexes can then be separated by SDS polyacrylamide gel elctrophoresis (Chapter 5).
The complete structure of membrane proteins is difficult. They do not crystallize well for X-ray crysallography (Chapter 5). This is bacteriorhodopsin. A small protein which acts as a membrane transport protein that pumps H+ out of the bacterium. It gets its energy from light, which is absorbed by retinal.Retinal changes shape
The structure of a bacterial photosynthetic reaction center includes four protein molecules.
The plasma membrane must be strengthened by the cell cortex. This is a framework of proteins attached to the membrane via transmembrane proteins.The shape and mechanical properties of the plasma membrane is determined by a meshwork of fibrous proteins - the cell cortex. Red blood cells are very simple, allowing study of the cell cortex in simple form. Genetic abnormalities in spectrin structure result in anemia, spherical, fragile rbcs
The spectrin-based cell cortex of human red blood cells. Much simpler than other cells. Dystrophin in muscular dystrophy
Most of the proteins in the plasma membrane have short chains of sugars (oligosaccharides) linked to them - glycoproteins. Others have longer polysaccharide chains - proteoglycans. All the glycoproteins, proteoglycans, and glycolipids are found on the noncytosolic side of the lipid membrane. They form a sugar coating called the glycocalyx. Glycocalyx halps to protect the cell surface from mechanical and chemical damage, absorb water and give the cell a slimy surface to help cells squeeze through narrow spaces and prevent them from sticking to each other or the walls of blood vessels.
Besides protection and lubrication, the glycocalyx is important in cell-cell recognition and adhesion. Some proteins (lectins) recognize particular oligosaccharide side chains and bind to them. The short oligosaccharides are enormously diverse, joined in different ways, branched, very complex and hard to study. Ex. Recognition of an egg by a sperm Specific carbohydrate chains on the surface of neutrophils binds a lectin on the cells of the blood vessels at the site of infection. Allowing them to stick transiently and then bind more strongly to other adhesion molecules. In this way the phagocytes enter and ingest the bacteria.
The lipid bilayer is a two-dimensional fluid. Many lipids and proteins move freely within the plane. This is demonstrated by staining mouse cells with rhodamine and human cells with fluorescein. These two cells are then fused. Within 30 minutes the proteins from the mouse and human cells have diffused and are intermixed.
However, lipids and proteins do not all float freely in the membrane. The cell controls the movement of many proteins. Cells have ways of confining particular plasma membrane proteins to localized areas, creating membrane domains which are functionally specialized. Proteins are moved together when signaled by receptors like adhesion molecules. Bound by the extracellular matrix Tethered to the cell cortex Stopped by diffusion barriors. Held by proteins on another cell
In epithelial cells that line the gut, uptake of nutrients from the gut is confined to the apical surface while proteins involved in the transport of solutes out into the tissues and bloodstream is confined to the basal and lateral surfaces. This asymmetric distribution is maintained by abarrier formed by tight junctions - seals between adjacent cells.