Download

1 / 25
400 likes | 846 Vues
Struttura e proprietà dei macronutrienti. La nutrizione potrebbe essere definita come l’utilizzazione dei cibi da parte degli organismi per la crescita, la riproduzione e il mantenimento della salute.

E N D
Struttura e proprietà dei macronutrienti
La nutrizione potrebbe essere definita come l’utilizzazione dei cibi da parte degli organismi per la crescita, la riproduzione e il mantenimento della salute. I composti classificati come nutrienti includono: acqua, carboidrati, proteine e aminoacidi, lipidi, vitamine e minerali. I nutrienti sono ottenuti dall’ingestione dei cibi e vengono utilizzati dal nostro corpo per l’ anabolismo, il metabolismo e il mantenimento dei tessuti.
I MACRONUTRIENTI organici sono composti del carbonio, questi includono: CARBOIDRATI, LIPIDI e PROTEINE. Come tutti i composti organici sono costituiti da sei elementi fondamentali: idrogeno, ossigeno, carbonio, azoto, zolfo e fosforo. Questi sei elementi con un peso atomico relativamente basso ( 32 ) costituiscono le strutture di proteine, lipidi e carboidrati, come anche di acidi nucleici e intermedi dei processi metabolici. G6P AcetilCoA
Se non si tiene in considerazione l’acqua, che fa parte del corpo umano per il 65%, carbonio, ossigeno, idrogeno e azoto per un uomo di 65 Kg costituiscono il 16% di proteine, il 18% di lipidi e solo lo 0,9% di carboidrati sottoforma di glicogeno.
La capacità del carbonio di formare legami C-C ottenendo lunghe catene e composti ciclici permette la formazione di una miriade di composti organici. Nelle molecole organiche, gli atomi di carbonio, idrogeno, azoto, zolfo e fosforo sono tenuti insieme da legami covalenti che si formano quando due elettroni condividono lo stesso orbitale esterno. Per la molecola organica ogni legame covalente rappresenta anche un piccolo deposito di energia.
I carboidrati, maggiormente presenti nella frutta, nella verdura, nei legumi e nei semi dei cereali sono responsabili del sapore e della consistenza degli alimenti; costituiscono la maggiore fonte di energia, soprattutto per gli esseri umani; sono digeriti e assorbiti nell’intestinotenue e in piccola parte dalla flora batterica intestinale localizzata nell’intestino crasso.
GLUCOSIO HO H Essenziale substrato energetico
Classificazione Le desinenze oso o osio indicano la presenza del gruppo aldeidico mentre uloso o ulosio, quelle dei gruppi chetonici. polidrossialdeidi (aldosi) e polidrossichetoni (chetosi) natura idrofilica Gruppi ossidrili Esteri Eteri Deossi Amino
Classificazione I monosaccaridi sono classificati in base al numero degli atomi di carbonio presenti nelle loro strutture, triosi, tetraosi, pentosi, esosi ed eptosi. In base alla loro stereoisomeria, D o L, e al grado di polimerizzazione, disaccaridi, oligosaccaridi e polisaccaridi. Da un punto di vista nutrizionale, il glucosio o D-aldoesoso monosaccaride è il più importante.
Le desinenze oso o osio indicano la presenza del gruppo aldeidico mentre uloso o ulosio, quelle dei gruppi chetonici. 2 1 ALDOSO CHETOSO Il gruppo carbonilico sul C-1 Il gruppo carbonilico sul C-2
Per gli zuccheri attivi nei sistemi biologici, lo stato D è fondamentale; nel caso del D-glucosio l’ossidrile, secondo le proiezioni di Fischer, si trova a destra del carbonio chirale. D-glucosio D-gliceraldeide L-glucosio
Le conformazioni a sedia e le proiezioni di Haworth sono le più usate per la rappresentazione degli zuccheri ciclici in biochimica.
I chetosi sono coinvolti come intermedi fosforilati nel metabolismo dei carboidrati.
L’acido N-Acetilmuramico, un chetoso acido a nove atomi di carbonio, è un importante segnale nelle glicoproteine. HO H OH HO OH OH HO
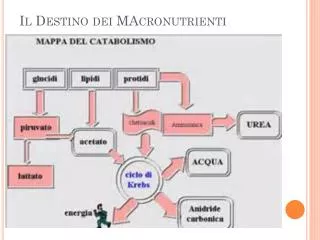
Metabolismo dei Carboidrati Tutti i monosi che compaiono sottoforma polimerica negli alimenti, prima di poter svolgere la loro funzione metabolica, devono essere convertiti in unità più piccole. Per la cellula la molecola del saccarosio come tale non è importante per ottenere energia. E’ la quantità di unità di glucosio presenti nel polisaccaride di cui la cellula ha bisogno. Le Amilasi e le Glucosidasi o Oligosaccaridasi sono gli enzimi capaci di scindere i polimeri dei carboidrati in unità singole.
Amilasi e Glucosidasi hanno diversa localizzazione Amilasi: bocca (ghiandole salivari) e intestino (pancreas-dotti) Glucosidasi: orletto a spazzola degli enterociti La digestione dei carboidrati inizia nella bocca sotto l’azione della a-amilasi che agisce sui legami glicosidici posizionati all’interno della catena polisaccaridica (granulo di amido). La trascrizione genica dell’enzima è influenzata da fattori cellulo-specifici nucleari come il fattore 1 nelle cellule del pancreas. La presenza di carboidrati nel lume intestinale e l’azione diretta della colecistochinina (ormone secreto dal duodeno e dal cervello) aumenta la sintesi di amilasi dalle cellule pancreatiche. La cottura dei cibi liberano i granuli di amido dalle proteine ad essi associati rendendo il polisaccaride più disponibile alla degradazione enzimatica.
Caratteristiche dell’a-amilasi Il pH ottimale per l’azione dell’enzima è quello della neutralità per cui la sua azione cessa quando il bolo alimentare viene deglutito e passa allo stomaco. Il pH acido dei succhi gastrici inattiva l’enzima, la degradazione dei carboidrati potrebbe continuare a pH acido ma richiederebbe molto tempo; lo stomaco si svuota prima che ciò possa avvenire. La presenza di amido nel pasto può comunque proteggere l’enzima dall’inattivazione da parte del pH 2, in questo modo l’enzima continua la scissione dell’amido nell’intestino. PREMATURI, presentano livelli molto bassi di amilasi pancreatica nel duodeno.
ATTIVITA’ ENZIMATICA DELL’ a-AMILASI L’enzima taglia dall’interno le catene polisaccaridiche legando consecutivamente 5 subunità oligosaccaridiche, il taglio è specifico ed avviene tra la seconda e la terza subunità del POLISACCARIDE; l’idrolisi interessa i legami a 1-4 glicosidici. sito di taglio dell’a-Amilasi 5 4 3 2 1 Amilosio sito attivo a-Amilasi si formano piccoli polimeri a 5 unità che verranno ulteriormente scissi nel trisaccaride maltotriosio e nel disaccaride maltosio.
ATTIVITA’ ENZIMATICA DELL’ a-AMILASI L’enzima continua ad avere la sua attività anche sulle catene polisaccaridiche con legami a 1-4 glicosidici che fanno parte dell’amilopectina, tuttavia la presenza di legami a 1-6 glicosidici portano alla formazione di a-destrine capaci di inibire l’enzima per l’idrolisi dei legami a 1-4; per questo motivo le destrine sono limitanti per l’azione dell’enzima. sito di taglio dell’a-Amilasi a 1-6 glicosidico a-destrina amilopectina maltosio maltotriosio
Orletto a spazzola L’orletto a spazzola degli enterociti dell’intestino tenue è ricoperto di speciali glicoproteine ad attività idrolasica. Responsabili dell’ultimo stage di idrolisi degli oligosaccaridi e dei disaccaridi. Le OLIGOSACCARIDASI sono espresse solo dagli enterociti, sono sintetizzate nel RE e trasportate all’orletto a spazzola grazie alla formazione di vescicole dall’apparato di Golgi. Questi enzimi hanno delle caratteristiche fondamentali.
OLIGOSACCARIDASI Da un punto di vista strutturale sono tutte costituite da regione N-terminale: Idrofobico che serve da sequenza segnale. Glicosilato. C-terminale: Ancoraggio alla membrana dell’orletto a spazzola La proteina è per il 95% orientata verso l’esterno della membrana cellulare come anche il sito catalitico. Maggiore efficienza Il corredo genico completo per la sintesi di queste glicoproteine è acquisito durante il periodo fetale
Oligosaccaridasi Le OLIGOSACCARIDASI si dividono in base al grado di specificità per il substrato. Glucoamilasi alta specificità per glucosiloligosaccaridi da 4 a 9 unità di glucosio, taglio dei legami a1-4 glicosidici. Sucrasi unico enzima che taglia i legami a1-b2 tra il glucosio e il fruttosio (saccarosio) effettua anche tagli a1-4 a-Destrinasi altaspecificità sui legami a1-6 delle destrine, taglio anche dei legami a1-4. L’enzima è chiamato anche isomaltasi Questi tre enzimi lavorano in maniera complementare gli uni rispetto agli altri per i tagli dei legami delle destrine per ottenere unità libere di glucosio.
Meccanismo di azione combinato dei tre enzimi glucosio a-destrina a destranasi/glucoamilasi glucosio sucrasi/glucoamilasi a destranasi sucrasi glucosio glucosio
Oligosaccaridasi Gli ultimi due enzimi che fanno parte delle OLIGOSACCARIDASI sono la Trealasi e la b-Galattosidasi Trealasi: scinde i legami a1-a1 presenti in alcuni disaccaridi contenenti due unità di glucosio b-Galattosidasi: presente dalla nascita, l’enzima ha alta specificità per il lattosio ed è indispensabile ai neonati per il metabolismo del latte materno. La quantità dell’enzima diminuisce con l’età.