Download

1 / 25
250 likes | 423 Vues
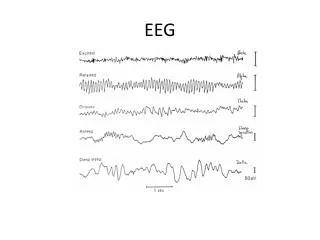
Rytmy EEG - pytania. Komórkowe i sieciowe mechanizmy generacji różnych wzorców w sygnałach EEG Znaczenie różnych rytmów w procesowaniu informacji Wstępujące i zstępujące układy neuromodulacyjne kontrolujące rytmy EEG Transformacja normalnych wzorców aktywności w wyładowania epileptyczne.

E N D
Rytmy EEG - pytania • Komórkowe i sieciowe mechanizmy generacji różnych wzorców w sygnałach EEG • Znaczenie różnych rytmów w procesowaniu informacji • Wstępujące i zstępujące układy neuromodulacyjne kontrolujące rytmy EEG • Transformacja normalnych wzorców aktywności w wyładowania epileptyczne David A. McCormick Section of Neurobiology Yale University School of Medicine http://www.med.yale.edu/neurobio/mccormick/seminars.html Thalamocortical Activity: Sleep, Wake, and Epilepsy
Generatory rytmów EEG – wrzeciona, kompleksy K i wolna oscylacja Wrzeciona snu składają się z dwóch rytmów. Rosnące i malejące wrzecionowate fale o częstości 7-14 Hz, trwające 1-2 sekundy, występujące z częstością wolnej oscylacji 0.2-0.5 Hz. Zapis u człowieka pochodzi z elektrod powierzchniowych podczas II fazy snu. Zapis u naturalnie śpiącego kota pochodzi z elektrod implantowanych.
Generatory rytmów EEG – wzgórze Wrzeciona generowane są we wzgórzu. Komórki przekaźnikowe wzgórza (Relay cells, Thalamocortical, TC) dostają wejścia sensoryczne (np. z siatkówki). Aksony komórek TC docierają do IV warstwy kory, a warstwa VI wysyła projekcje zwrotne do wzgórza, co tworzy pętlę korowo-wzgórzową. Komórki siatkowate wzgórza (nucleus Reticularis thalami, nRT, Reticular cells, RE) tworzące ‘płaszcz’ otaczający wzgórze, są GABAergiczne i unerwiają gęsto komórki TC. W ten sposób zapewniają hamowanie ‘do przodu’ (feedforward) na drodze kora-wzgórze i hamowanie zwrotne (feedback) pomiędzy TC i RE. Wzgórze i kora są pod kontrolą różnych układów modulujących m.in. cholinergicznego (ACh), noradrenergicznego (NE), serotoninergicznego (5-HT), histaminergicznego (HA).
Badania in vitro Komora in vitro pozwalająca na zapisy wewnątrzkomórkowe ze skrawków mózgu. Manipulator z prawej strony służy do utrzymania elektrod, manipulatory z lewej strony służą do aplikacji leków. Skrawki są utrzymywane w temperaturze 35oC przez 12 godzin. Skrawki mózgu fretki
Dwa rodzaje aktywności komórek wzgórza Zapisy wewnątrzkomórkowe z komórek TC ukazujące dwa rodzaje aktywności. Niektóre komórki generują rytmiczne salwy potencjałów czynnościowych. Wstrzyknięcie prądu depolaryzującego zamienia salwy w pojedyncze potencjały czynnościowe. Usunięcie prądu przywraca wolny rytm. Podobne własności wykazują komórki RE. Zmiany na poziomie pojedynczego neuronu odzwierciedlają zmiany w EEG – rytmiczne salwy podczas rytmów snu i aktywność toniczna w czasie czuwania.
Oddziaływanie TC - RE Pojedyncza komórka TC w ciele kolankowatym bocznym. Kolaterale aksonu dochodzą do jądra siatkowatego, a akson główny początkuje drogę do pierwszorzędowej kory wzrokowej. Równoczesne zapisy wewnątrzkomórkowe z neuronów TC i RE pokazują, że salwa potencjałów czynnościowych w pojedynczej komórce RE wywołuje hamujące potencjały postsynaptyczne w komórkach TC, mogące wywołać salwę po wyjściu z hiperpolaryzacji (E) . Pojedyncza komórka RE w ciele okołokolankowatym. Aksony komórek RE unerwiają wyłącznie komórki TC.
Oddziaływanie TC – RE cd Salwa potencjałów czynnościowych w komórce RE wywołuje hamujące potencjały postsynaptyczne (IPSP) w komórce TC (LGNd), mogące wywołać salwę po wyjściu z hiperpolaryzacji (rysunek D). Salwa w komórce TC wywołuje serię pobudzających potencjałów postsynaptycznych (EPSP) w komórce RE (PGN) (rysunek H). Salwa w komórkach RE, inicjuje kolejny cykl oscylacji. Dwusynaptyczny obwód jest generatorem wrzecion in vitro.
Generacja wrzecion i wyładowań epileptycznych W doświadczeniach in vitro, wrzeciona generowane są poprzez rytmiczne oddziaływanie komórek RE (PGN) z komórkami TC ( LGNd). Izolowane komórki jądra siatkowatego wzgórza (RE) nie są w stanie generować rytmicznej aktywności o częstości wrzecion. Blokada receptorów GABAA prowadzi do transformacji wrzecion w wyładowania przypominające epileptyczne napady nieświadomości. A. potencjał polowy we wzgórzu. W sygnale przefiltrowanym widoczne wrzeciona oraz wolna oscylacja. B. generacja wrzecion in vitro w pętli RE – TC. Komórki RE hiperpolaryzują dużą liczbę komórek TC. Komórki TC wychodząc z hiperpolaryzacji odpalają salwę potencjałów czynnościowych, która aktywuje komórki RE i powoduje u nich odpalanie salwy i inicjacje kolejnego cyklu. C. Blokada receptorów GABAA powoduje dezinhibicję komórek RE i ich silniejsze wyładowania. Dłuższe salwy RE aktywują receptory GABAB co prowadzi do wydłużenia hiperpolaryzacji, silniejszych salw w komórkach TC i synchronizacji aktywności.
Transformacja wrzecion w wyładowania epileptyczne in vivo Systematyczna aplikacja penicyliny (bloker receptora GABAA) do kory kota prowadzi do transformacji wrzecion w napady epileptyczne typu ‘iglica i fala’ (spike-wave). Górny sygnał jest potencjałem polowym, dolny wykres ukazuje autokorelogram. Przy każdym rysunku podany jest czas od początku aplikacji penicyliny. Transformacja wrzecion w aktywność epileptyczną w wyniku rozległej aplikacji penicyliny do kory kota. Przed aplikacją stymulacja nucleus centralis medialis (NCM) prowadzi do opowiedzi normalnej, po aplikacji – do patologicznej.
Generator wrzecion in vivo – jądro siatkowate wzgórza In vivo generatorem wrzecion jest jądro siatkowate wzgórza. Komórki RE są hamujące i są połączone zarówno poprzez aksony, jak i złącza dendro-dendrytyczne. Hipoteza generatora RE opiera się na: 1. Jądra RE pozbawione wejść generują wrzeciona oraz wolną oscylację. 2. Wrzeciona we wzgórzu i korze zanikają po przecięciu połączeń z RE. Zapis górny: potencjał polowy w jądrze siatkowatym wzgórza (RE) po odcięciu połączeń z korą, wzgórzem i pniem mózgu. Widoczne wrzeciona oraz wolna oscylacja. Poniżej: zapisy EEG z kory po przecięciu połączeń kory ze wzgórzem. Widoczne fale delta, lecz wrzeciona są nieobecne.
Generator wrzecion in vivo – jądro siatkowate wzgórza Próba rozwikłania paradoksu istnienia różnych generatorów wrzecion in vivo i in vitro została podjęta z pomocą modelu komputerowego. Gdy potencjał spoczynkowy komórek RE w modelu odpowiadał sytuacji in vitro (-65 mV – -85 mV), komórki nie były w stanie podtrzymać aktywności gdyż potencjał spoczynkowy był zbyt blisko potencjału równowagi dla receptora GABAA(~ -80 mV). Depolaryzacja komórek RE do potencjału odpowiadającego sytuacji in vivo (-60 mV – -70 mV) wywołuje oscylacje o częstości wrzecion. Depolaryzacja in vivo (symulowana w modelu) jest spowodowana działaniem neuromodulatorów – serotoniny i noradrenaliny. Symulacje komputerowe połączonych komórek RE. Oddziaływania pomiędzy dwoma komórkami RE generuje aktywność rytmiczną z przedziału 6-9 Hz. Symulacje komputerowe sieci 100 połączonych komórek RE. Pokazane zapisy z 10 komórek oraz ich uśredniony potencjał błonowy. W wyniku aktywacji 20% synaps NE/5HT (noradranaliny/serotoniny) potencjał oscyluje w częstości 10-16 Hz. Wyłączenie aktywacji prowadzi do hiperpolaryzacji i zaniku aktywności, co może odpowiadać sytuacji in vitro.
Generator wrzecion – pełny obwód RE - TC - Cx W nienaruszonym obwodzie RE-TC-Cx, komórki TC przyczyniają się do generacji wrzecion. Świadczy o tym m.in. fakt, że długość hyperpolaryzacji w komórkach TC ma wpływ na częstość wrzecion. Bez komórek TC, wrzeciona nie byłyby widoczne w powierzchniowych zapisach EEG. Góra: obwód trzech rodzajów komórek: RE komórki jądra siatkowatego wzgórza (reticular thalamic nucleus), TH-cx (komórki wzgórzowo-korowe (thalamocortical), Cx komórki kory (cortex). Zapis EEG ukazuje wrzeciona i wolna oscylację. Poniżej: zapisy wewnątrzkomórkowe u uśpionych kotów. W trakcie generacji wrzeciona komórki TC odpalają salwy tylko w niektórych cyklach.
Generator wrzecion – pełny obwód RE - TC - Cx Rola komórek kory polega na czasowo-przestrzennej synchronizacji indywidualnych generatorów wrzecion we wzgórzu. Czasowo-przestrzenne mapy aktywności we wzgórzu. Strzałka wskazuje 1s. Każda kolumna reprezentuje 8 mm (8 elektrod) wzgórza, a kolor wskazuje napięcie lokalnego potencjału polowego. Po usunięciu kory mózgowej kota, aktywność wrzecion jest zachowana lecz ich synchronizacja maleje. Dolne panele pokazują korelację aktywności dla wszystkich możliwych par elektrod in vivo i w symulacji komputerowej
Korowe napady nieświadomości Podczas napadów nieświadomości u kota, komórki TC we wzgórzu są w stanie hieperpolaryzacji i nie wykazują aktywności. Wskazuje to na korowy generator napadów. Zapis pierwszy: napad nieświadomości u kota manifestujący się aktywnością ‘iglica-fala’ w EEG. Zapis drugi: aktywność komórki w korze motorycznej. Zapis trzeci: aktywność neuronu wzgórza. W trakcie napadu komórka wzgórza wykazuje toniczną hiperpolaryzację, z której wychodzi po zakończeniu napadu.
Model - ‘wzgórzowe’ napady nieświadomości Zablokowanie receptora GABAA we wzgórzu powoduje zmianę częstości oscylacji z 11 na 4 Hz i wydłużenie salw w komórkach RE oraz TC. W komórkach kory (PY i IN) następuje jedynie zmiana częstości. Potencjał polowy kory nie przypomina ‘iglicy i fali’.
Model - ‘korowe’ napady nieświadomości Zablokowanie receptora GABAA we korze prowadzi do transformacji wrzecion w synchroniczną aktywność ‘iglica i fala’ o częstości 3 Hz.
Generatory rytmów EEG – wolna oscylacja Wolna oscylacja – rytm o częstości < 1 Hz A. Wolna oscylacja (~ 0.9 Hz) u kota widoczna w powierzchniowym i głębinowym EEG oraz zapisy wewnątrzkomórkowe z neuronu w korze motorycznej (obszar 4) i z neuronu wzgórza. W części B widać powiększenia cyklu. Maksimum w EEG odpowiada potencjałom czynnościowym w komórkach korowych. Po maksimum występuje krótka sekwencja wrzeciona w EEG. Komórka wzgórza generuje salwę z opóźnieniem 150-200 ms w stosunku do aktywności komórek kory.
Generatory rytmów EEG – wolna oscylacja cd Wolna oscylacja jest zjawiskiem sieciowym (generatorem jest sieć neuronów kory, a nie pojedyncze komórki lub lokalny obwód). Świadczą o tym: a) występowanie rytmu w korze, po usunięciu wzgórza, b) zanik rytmu we wzgórzu po usunięciu kory, c) zakłócenie rytmu po zablokowaniu połączeń wewnątrz-korowych Zapisy wewnątrzkomórkowe oraz sygnały EEG z dwóch obszarów położonych w przeciwnych półkulach. A, B – dwie różne pary zapisów. W obu przypadkach, wolna oscylacja w EEG pojawia się dopiero gdy obie komórki zaczynają jednocześnie wykazywać głębokie hiperpolaryzacje.
Generatory rytmów EEG – wolna oscylacja cd Dezorganizacja wolnej oscylacji w wyniku zablokowania synaps wewnątrz-korowych. Wykresy ukazują sekwencyjne korelacje wzajemne (cross-correlation) zapisów wewnątrzkomórkowych z przedniej i tylnej części mózgu kota. Na schemacie mózgu widać położenie elektrod i miejsce aplikacji lidocainy (strzałka). Po aplikacji korelacje wzajemne maleją i tracą wyraźne centralne maksimum – świadczy to o desynchronizacji aktywności.
Generatory rytmów EEG – wolna oscylacja cd Zapisy wewnątrzkomórkowe podczas wolnej oscylacji z komórek kory i wzgórza. Górny zapis - wolna oscylacja (0.3 Hz) w neuronie korowym. Krótkotrwała stymulacja włókien cholinergicznych (pozioma linia - PPT) powoduje zanik oscylacji i aktywność toniczną. Poniżej: dwa neurony korowe wykazujące rytm 0.17 Hz i 0.3 Hz. W jednym z zapisów widoczna aktywność delta (1.5 Hz) generowana w komórkach Th-cx. Komórka RE wykazuje wolną oscylację (0.3 Hz), komórka Th-cx wykazuje aktywność delta (2.5 Hz) oraz wolny rytm (0.2-0.3 Hz).
Generatory rytmów EEG – kompleksy-K Zespół grafoelementów składający się z ostrej iglicy, wolnej fali i wrzeciona nazywa się kompleksem-K. Zapisy głębinowe (EEG) i wewnątrzkomórkowe z komórek TC i RE ukazujące ostrą iglicę, wolną falę i oscylacje wrzecionowatą.
Wolna oscylacja grupuje rytmy EEG snu Wolna oscylacja łączy rytmy korowe w czasie snu (wrzeciona, kompleksy-K i fale delta) A. Kompleksy-K w zapisie 3 fazy snu u człowieka. Oba kompleksy K są ‘umieszczone’ w wolnej oscylacji (~0.6 Hz). B. Uśredniony kompleks-K. C. Widmo mocy z elektrody C3. Trzy zakresy częstości zaznaczone zaznaczone są w okienku oraz w części D rysunku.
Generatory rytmów EEG – fale delta Fala delta jest rytmem o częstości 1- 4 Hz występującym w 3 i 4 fazie snu. Istnieją dwa rodzaj fal delta – jeden z nich generowany jest w korze i nie zanika po usunięciu wzgórza (mechanizm nieznany). Drugi rodzaj fal delta generowany jest we wzgórzu i nie zanika po usunięciu kory (mechanizm dobrze poznany). Fale delta (3 - 4 Hz) pogrupowane wolną oscylacją (0.3 – 0.4 Hz). A. Zapis z komórki wzgórza. B. Od góry do dołu: zapis zewnątrzkomórkowy z komórki kory, lokalny potencjał polowy i zapis EEG. Kawałki sygnału oznaczone ‘*’ są pokazane w powiększeniu poniżej. C. Autokorelogramy dwóch komórek korowych. Obie komórki wykazują wolną oscylacje, a komórka b dodatkowo wykazuje rytm delta.
Generatory rytmów EEG – fale delta cd Wzgórzowa fala delta jest generowana w komórkach TC w wyniku oddziaływania prądu wapniowego IT i prądu Ih aktywowanego hiperporalyzcją. -Aktywacja prądu IT depolaryzuje błonę powyżej progu prądów Na+/K+. -Depolaryzacja dezaktywuje prąd Ih aktywnego przed wystąpieniem potencjału wapniowego -Repolaryzacja błony jest wynikiem dezaktywacji prądu IT. Głęboka depolaryzacja jest wynikiem zmniejszonego wpływu prądu depolaryzującego Ih. -Hiperpolaryzacja de-inaktywuje (= usuwa inaktywację) prąd IT oraz aktywuje prąd Ih, co prowadzi do kolejnej depolaryzacji. A. Generacja fali delta w komórkach TC. B. Oscylacje delta w komórkach TC wywołane prądem hiperpolaryzującym. Im większa hiperpolaryzacja (od góry do dołu), tym więcej cykli fal delta. Prąd depolaryzujący (strzałki) zatrzymuje oscylację, a jego usunięcie ją przywraca.
Wrzeciona i fale delta – zależność od potencjału błonowego w TC Potencjał błonowy komórek TC odpowiada za przejście od wrzecion do fal delta. A-C ukazują zapisy wewnątrzkomórkowe komórki TC i EEG z obszaru kory motorycznej u kota. A. Desynchronizacja w EEG jest związana z aktywnością toniczną komórki. B. Hiperpolaryzacja komórki prowadzi do pojawienia się wrzecion (8 -10 Hz) w EEG pogrupowanych wolną oscylacją (~ 0.8 Hz). C. Podanie prądu hiperpolaryzującego (-0.2 nA) do komórki zmienia wrzeciona w fale delta (3 - 4 Hz) również pogrupowane wolną oscylacją (~ 0.8 Hz).