Download
1 / 42
420 likes | 552 Vues
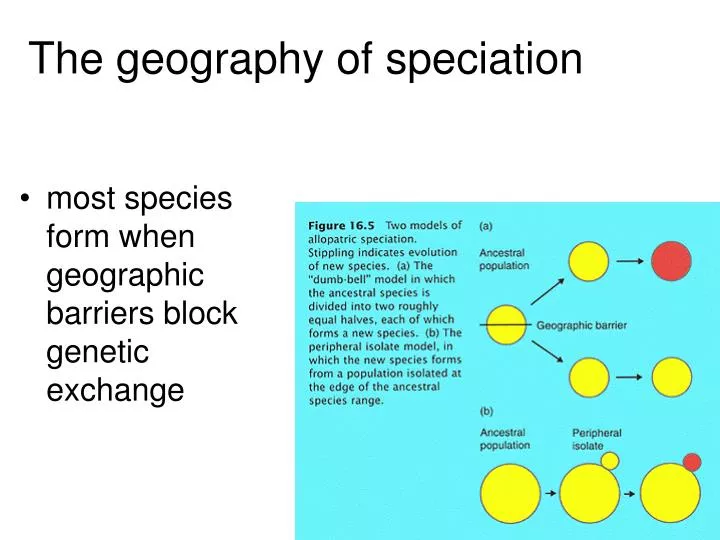
The geography of speciation. most species form when geographic barriers block genetic exchange. Allopatric speciation. arguably the most common way species form evidence for allopatric speciation is common in biogeography: related species often occupy nearby, non-overlapping ranges.

E N D
The geography of speciation most species form when geographic barriers block genetic exchange
Allopatric speciation arguably the most common way species form evidence for allopatric speciation is common in biogeography: related species often occupy nearby, non-overlapping ranges
Allopatric speciation Isthmus of Panama closed ~ 3.1 MYA • Split ~150 “geminate” (twin) species
Evidence for allopatric speciation in snapping shrimps Knowlton et al.(1993) created a phylogeny of Pacific (P) and Carribean (C) species pairs of Alpheus In 6 out of 7 cases, the closest relative of a species was on the other side of the Isthmus
Allopatric speciation through “dispersal and colonization”
Evidence from phylogeny of Hawaiian Drosophila D. heteroneura D. silvestris
Founder effect speciation divergence of a small population isolated on periphery of range • thought to explain species radiations on islands
Founder effect speciation the genetic “founder effect” of small population size itself is thought to drive speciation
Sympatric speciation no geographic barrier much rarer but shows how ecology can drive speciation by selecting for assortative mating from Barluenga et al. (2006) Nature 439: 719-723
Sympatric speciation valid cases: cichlids in crater lakes host races of Rhagoletis (apple maggot fly) from Barluenga et al. (2006) Nature 439: 719-723
Lake Apoyo: an isolated volcanic crater lake, homogeneous habitat < 23,000 years old
mtDNA sequences are monophyletic: no Lake Apoyo sequences are found in sister species outside the Lake This shows that all Lake Apoyo fish evolved in sympatry
Morphological and ecological differences evolved sympatrically benthic limnetic
Sympatric speciation in action Rhagoletis pomonella: the apple maggot fly Larvae feed on natural host: hawthorn fruits A “host race” infesting apple trees appeared in North America ~150 years ago apples hawthorns
apples hawthorns
Speciation in progress the apple race and hawthorn races have evolved reproductive barriers Adults mate and oviposit on their host trees This leads to limited interbreeding
Speciation in progress the apple race and hawthorn races have evolved reproductive barriers Adults mate and oviposit on their host trees Genetic differences between the races mark the early stages of speciation
apples hawthorns Genetic differences between apple and hawthorn races • the two races show genetic differences at six allozyme loci • included is an aconitase locus called Acon-2 citrate aconitate isocitrate
apples hawthorns Genetic differences between apple and hawthorn races • the frequency of the Acon-2 95 allele is much higher in the hawthorn race
The role of natural selection in divergence of Rhagoletis • Feder and coworkers predicted that natural selection opposes migration and creates allozyme differences • One hypothesis: hawthorns ripen fruits 3-4 weeks later than apples • Apple fly larvae experience longer periods of warm temperatures prior to and while pupating • This must select for divergent genes
Their experiment... • Collect hawthorn pupae and expose to warm temperatures for a varying duration • Follow with freezing temperatures (“winter”), then by warming (“spring”) • Collect emerging adults and assay allozymes
hawthorn pupae exposed to longer periods of “pre-winter” warmth produced adults with allele frequencies approaching those of the apple race
The classic “three stage” model of allopatric speciation • 1st stage: a geographic barrier creates isolation between two or more portions of a population • 2nd stage: the descendant populations diverge genetically (due to drift and selection) [Rhagoletis has skipped the 1st and is in the 2nd stage]
“Ecological speciation” in sticklebacks: more evidence for natural selection during the 2nd stage • Dolph Schluter and coworkers have studied Gasterosteus aculeatus in rivers and lakes of British Columbia for many years • A marine ancestor colonized rivers at least 3 times, independently
“Ecological speciation” in sticklebacks shows evidence for natural selection during the 2nd stage • Several lake colonizations have led to independent cases of “ecological speciation”
“Ecological speciation” in sticklebacks shows evidence for natural selection during the 2nd stage • In several lakes, two morphs that show differences in diet, morphology, and behavior have diverged: • A smaller limnetic (open water) form • A larger benthic (bottom dwelling) form • Benthic and limnetic forms mate like-with-like
Evidence for a role for sexual selection during the “divergence” stage
Sexual selection on head width in D. heteroneura Males with wider heads are chosen by females on leks And they win contests with males for territories
This means that sexual selection is likely to be responsible for the differences in head shape between D. heteroneura and D. silvestris And that it served a role in their speciation??
The classic “three stage” model of allopatric speciation • 3rd stage: reproductive isolation is completed, or perfected • this occurs after “secondary contact” between allopatric populations • a crucial step, why? • secondary contact is common • without complete reproductive barriers, species will re-fuse
Secondary contact after Pleistocene glaciations: bird hybrid zones from Futuyma (1998), p. 258
Secondary contact: hybrid zones in blue mussels Hilbish et al. 2000
Pacific mussels reinvaded the North Atlantic during warm, high sea level period about 15-20,000 ya from Riginos and Cunningham 2005
The classic “three stage” model of allopatric speciation • Reproductive isolation can be completed in two distinct ways • As a byproduct of the divergence process, through drift and selection (unrelated to interbreeding) • Via selection against hybridization (reinforcement)
Coyne and Orr’s (1997) survey of Drosophila species pairs • Reviewed data on over 150 species pairs • Whether the species are allopatric or sympatric • Genetic distance between the species pair (based on allozymes) as an estimate of age • The amount of postzygotic and prezygotic isolation
Their results... Prezygotic isolation increases with genetic distance Genes for prezygotic isolation diverge over time, just like the rest of the genome
Full isolation evolves in allopatry Secondary contact is not necessary to complete the speciation process
Coyne and Orr’s (1997) survey supports reinforcement Prezygotic isolation evolves faster in sympatry Selection against hybridization drives more rapid evolution
This is not “sympatric speciation!” why not?