Download

1 / 13
140 likes | 178 Vues
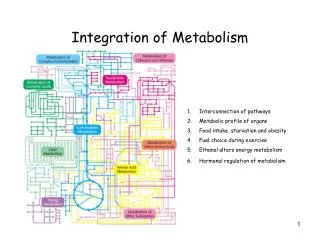
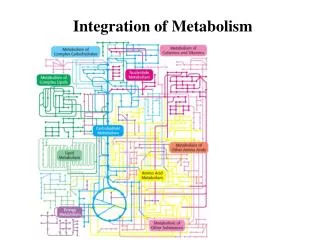
Integration of Metabolism. Metabolic pathways involved in energy production Glycolysis, gluconeogenesis, TCA cycle, oxidative phosphorylation, fat synthesis and breakdown Carbohydrate vs. fat vs. protein Molecules at the crossroads Regulation of glycogen storage and breakdown

E N D
Integration of Metabolism • Metabolic pathways involved in energy production • Glycolysis, gluconeogenesis, TCA cycle, oxidativephosphorylation, fat synthesis and breakdown • Carbohydrate vs. fat vs. protein • Molecules at the crossroads • Regulation of glycogen storage and breakdown • Regulation of glycolysis vs. gluconeogenesis • Insulin and Glucagon • Organ Specialization Lightning review of Biochemistry 463 Jason Kahn: Integration of Metabolism
Overview of Metabolic Regulation • We need to understand how metabolic regulation is supposed to work before we can understand how it fails in diabetes. • See Voet and Voet 3/e, Chapter 27 and chapters 19-26. Magisterial but dense. • Flux through pathways is determined at a few key steps. • Levels of critical metabolites are highly regulated, largely allosterically: much more rapid than gene expression. • Blood glucose level is the critical mis-regulated metabolite in diabetes. Many downstream consequences! Jason Kahn: Integration of Metabolism
Glycolysis • V+V 3/e chapter 17 • Conversion of Glucose to 2 Pyruvate: partial oxidation of glucose • Anaerobic and fast! Provides ATP for intense short term muscle activity • Generates 2ATP and 2 NADH (reducing power) • Net reaction:Glucose + 2 NAD+ + 2ADP + 2Pi 2NADH + 2 pyruvate + 2ATP + 2 H2O + 4H+ • Pyruvate is a key branch point metabolite. Jason Kahn: Integration of Metabolism
The Reactions of Glycolysis • V+V chapter 17 • First step is phosphorylation of glucose to give glucose-6-phosphate (G6P): keep expensive metabolites from escaping! • G6P is the connection to glycogen storage and breakdown pathways and the pentose phosphate pathway (which oxidizes glucose to provide biosynthetic reducing power in the form of NADPH). • 3rd step is fructose-6-phosphate to fructose-1,6-bisphosphate: substrate level phosphorylation catalyzed by phosphofructokinase I. This is the key regulated step. • In general, always exert flux control at irreversible (inelastic) steps, preferably early in the pathway to avoid unnecessary buildup of metabolites. To control the flux of a reversible (elastic) step is much more difficult, because the rates of forward and reverse reactions are both large, so flux adapts rapidly to substrate concentration. • Overall reaction is strongly downhill. Jason Kahn: Integration of Metabolism
The Fate of Pyruvate • In order to maintain a steady state, NADH produced in glycolysis must somehow be reoxidized to NAD+. • Under aerobic conditions, NADH is typically reoxidized to NAD+ in the process of oxidative phosphorylation to power phosphorylation of ADP to give ATP. This provides much more ATP than is directly produced in glycolysis: the Pasteur effect. • In this case, pyruvate is decarboxylated to give acetyl-CoA, with accompanying reduction of an additional NAD+ to NADH. Acetyl-CoA feeds into either the TCA cycle or fatty acid synthesis • Under anaerobic conditions, NAD+ must be regenerated by fermentation. • In yeast, alcoholic fermentation converts pyruvate to EtOH and CO2, via decarboxylation of pyruvate to acetaldehyde and then reduction of acetaldehyde to ethanol, which oxidizes NADH. (EtOH also poisons the yeast’s competition.) Consumption of ethanol produces NADH. • During vigorous exercise, the rate of oxidative phosphorylation in musclemitochondria cannot keep up with demand for ATP, so NADH builds up. NAD+ is regenerated by homolactic fermentation, the reduction of pyruvateby NADH to give lactate. The lactate buildup and pH drop cause musclefatigue (“the burn”). The lactate is shipped off to the liver, where it is convertedback to glucose (see below) • Ethanol consumption will convert pyruvate to lactate and prevent gluconeogenesis. Yet another reason not to drink on an empty stomach! Jason Kahn: Integration of Metabolism
Acetyl-CoA • Pyruvate dehydrogenase decarboxylatespyruvate to acetyl-CoA. Like alldecarboxylations, the reaction is irreversible: • The high-energy thioester bond of acetyl-CoAallows it to act as a carrier of activated acetylgroups. • It is the control point for deciding whether fluxwill be directed toward production of energyfor current requirements (the TCA cycle andoxidative phosphorylation) or toward storageof energy for the future (fatty acid synthesis). • The oxidative metabolism of fat feeds in toenergy production by providing NADH duringß-oxidation and then acetyl-CoA metabolism.Since acetyl-CoA cannot be converteddirectly to pyruvate, and its carbons are lost as CO2 ingoing through the citric acid cycle, stored fat cannot beconverted to sugar (unless one happens to be a plantand has access to the glyoxylate cycle). • Therefore, if one is living on stored fat rather than carbohydrates (i.e. one is starving or diabetic), acetyl-CoA is converted to ketone bodies (acetoacetate and hydroxybutarate) in the liver (ketogenesis) in order to feed the brain, heart and other organs. Spontaneous decarboxylation of acetoacetate gives acetone. This is why the breath of a person with uncontrolled diabetes can smell like acetone, and ketoacidosis or ketosis also causes dehydration and kidney damage. Jason Kahn: Integration of Metabolism
The TCA cycle • The TCA cycle is the metabolic hub of the cell. Occurs in the mitochondria. • Citrate synthase adds the two-carbon acetyl-CoA to oxaloacetate to form citrate (a tricarboxylic acid) • A series of reactions includes two decarboxylation steps in converting citrate back to oxaloacetate • Thus the carbon that started in glucose (or fat) has been completely oxidized to CO2, with the electrons used to reduce NAD+ and FAD to NADH and FADH2. • The cycle does not permit net incorporation of carbon from fat into oxaloacetate, which is the feedstock for gluconeogenesis: fat cannot be turned into sugar! • Most amino acid catabolism feeds into the TCA cycle as well: protein can be converted intosugar (and hence fat). • Largely regulated by feedback inhibition, the availability of substrate acetyl-CoA, and the NADH/NAD+ ratio, which in turn depends on oxidative phosphorylation and the cellular ATP/ADP ratio. • Ox-phos is the coupling of ATP synthesis with electron transfer from NADH to O2, which completes the combustion of glucose. Jason Kahn: Integration of Metabolism
TCA cycle II Metabolic Hub Product inhibition by NADH Jason Kahn: Integration of Metabolism
Gluconeogenesis • The body (mainly the liver) does not store enough glucose or glycogen to feed the brain for more than about 12 hr (the brain is a picky eater!). • Gluconeogenesis is the synthesis of glucose from pyruvate or oxaloacetate, hence the reverse of glycolysis. It is necessary for buffering blood glucose levels during fasting. • Since glycolysis is strongly downhill, gluconeogenesis cannot be the microscopic reversal of glycolysis: the three irreversible steps of glycolysis are driven by phosphate ester hydrolysis in the reverse reactions (red). The reactions in blue can go both ways. • The red reactions are carried out by different enzymes in the two directions, and are the control points for the glycolysis vs. gluconeogenesis decision. Fructose bisphosphatase and glucose-6-phosphatase are regulated by allostery, PEPCK is transcriptionally regulated, by glucocorticoids among others. • Pyruvate carboxylase + phosphoenol pyruvate carboxy kinase (PEPCK) reversing the last irreversible step of glycolysis. Jason Kahn: Integration of Metabolism
Gluconeogenesis II • Obviously, doing glycolysis and gluconeogenesis in the same tissue at the same time would be a bad idea, a “futile cycle” producing only heat. • However, in the Cori cycle, gluconeogenesis in the liver supports the muscle’s ability to produce ATP very rapidly through glycolysis and homolactic fermentation. • This means the liver must subsequently replace the ATP used up in gluconeogenesis, which means that oxidative phosphorylation is upregulated: the consequent increase in oxygen usage is “oxygen debt.” • Demonstrates that opposing pathways can occur at the same time as long as they aren’t in the same cell or compartment. Often the liver is generating glucose for export at the same time as the brain and muscle are consuming it. Jason Kahn: Integration of Metabolism
Glucose vs. glycogen vs. fat • Fat is roughly (CH2)n. The oxidation state of the carbon is –2. Fat is formed from excess carbon (acetyl-CoA) and reducing power (NADPH). • Carbohydrate is C(H2O)n, so the oxidation state of carbon is 0. • Hence fat is more reduced than carbohydrate, and provides more energy upon oxidation to CO2. (9 kcal/g vs 4). • It’s like the difference between burning wood vs. gasoline. • Fat is also be stored in a dehydrated form in adipose tissue, whereas sugars are hydrated, therefore less dense. • Glycogen is a polymer of glucose, a storage form that does not increase osmotic pressure as free glucose would. Stored in granules in muscle and liver. • Glucose is mobilized very rapidly through high-flux glycolytic pathway. Glycogen must be broken down by glycogen phosphorylase first, therefore more slowly metabolized. Fat must go through the relatively slow TCA cycle and oxidative phosphorylation to provide energy. Fat cannot be metabolized anaerobically. • Bottom line: the denser the energy storage, the more slowly it can be used. • Glucose in excess of current metabolic needs is converted to glycogen or fat, which are later broken down to either replenish glucose or drive ATP synthesis directly. Jason Kahn: Integration of Metabolism
Glycogen Metabolism • Glycogen is a polymeric form of glucose, with (14) linkages and (16) branches. Similar to starch. Cellulose has (14) linkages, giving sheet structures that are much harder to break down. Primarily found in muscle and liver. • Glycogen is synthesized by glycogen synthase, from glucose-6-phosphate via phosphoglucomutase and UDP-glucose pyrophosphorylase. Glycogen synthesis is energetically uphill. • Glycogen is broken down by glycogen phosphorylase and debranching enzyme; this is exergonic in the cell. It is degraded starting from an end, so the branched structure means that glucose can be released rapidly. Debranching is slower. • Glycogen synthesis and are catalyzed by different enzymes, so they can be independently regulated and do not occur simultaneously in the same tissue. Jason Kahn: Integration of Metabolism
Insulin and Glucagon • Insulin: Signals fed state. • Stimulates glucose uptake by muscle and other tissues. • Stimulates glycogen synthesis and fatty acid synthesis. • Represses gluconeogenesis and glycogen breakdown. • Upregulates genes for fatty acid biosynthesis. • Acts through receptor tyrosine kinase via many pathways in the cell. • Glucagon: Signals hungry state. • Represses glycogen synthesis and fatty acid synthesis. • Stimulates gluconeogenesis and glycogen breakdown, leading to glucose export by the liver. • Upregulates gene for PEPCK. • Acts through G-protein coupled receptor that activates adenylate cyclase. • Diabetes is caused by either lack of insulin (Type I) or lack of proper response (Type II), leading to increased blood glucose because uptake is not turned on. Jason Kahn: Integration of Metabolism