Download
1 / 75
800 likes | 1.32k Vues
Eduard Buchner (1860-1917) 1897 found fermentation in broken yeast cells 1907 Nobel Prize in Chemistry. The whole pathway in yeast and muscle cell were elucidated by. Arthur Harden 1865-1940. Glycolysis.

E N D
Eduard Buchner (1860-1917) 1897 found fermentation in broken yeast cells 1907 Nobel Prize in Chemistry
The whole pathway in yeast and muscle cell were elucidated by Arthur Harden 1865-1940
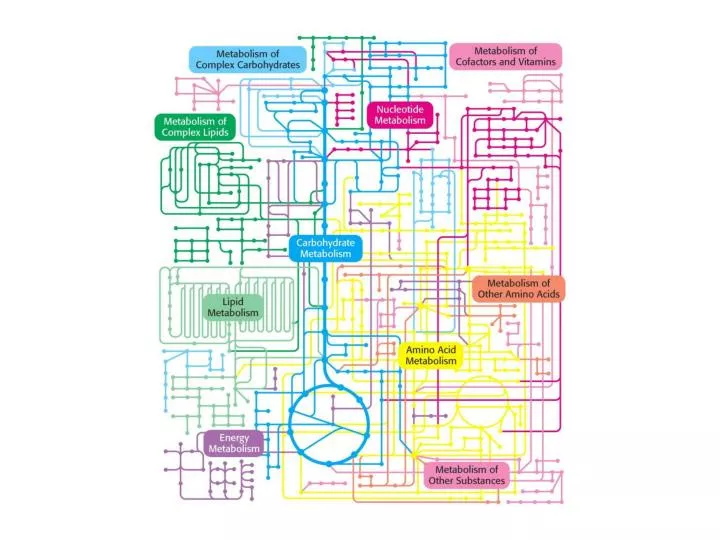
Glycolysis • Glycolysis is an almost universal central pathway of glucose catabolism, the pathway with the largest flux of carbon in most cells. • In some mammalian tissues (erythrocytes, renal medulla, brain, sperm), the glycolytic breakdown of glucose is the sole source of metabolic energy.
Glycolysis • Some of the starch-storing tissues, like potato tubers, and some aquatic plants derive most of their energy from glycolysis. • Many anaerobic microorganisms are entirely dependent on glycolysis.
Phosphohexose isomerase reaction by an active-site His residue Glu
3. Phosphorylation of fructose 6-phosphate: the first committed step in glycolysis
PFK-1 is named so because there is another enzyme catalyzes a similar reaction
In some bacteria, protists and (all) plants, a pyrophosphate-dependent phosphofructokinase (PFP) also catalyzes this reaction in a reversible way
Class I aldolases form Schiff base intermediate during sugar cleavage reaction • Class I aldolases were found in animals and plants. • Class II aldolases (fungi and bacteria) do not form the Schiff base and require a zinc ion to catalyze reaction.
Dihydroxyacetone phosphate and glyceraldehyde 3-phosphate become indistinguishable after triose phosphate isomerase reaction
6. Oxidation of glyceraldehyde 3-phosphate to 1,3-bisphosphoglycerate
The glyceraldehyde 3-phosphate dehydrogenase reaction Heavy metal ion such as Hg2+ will react with Cys residue, hence irreversibly inhibits the enzyme. hemiacetal
Glyceraldehyde 3-phosphate dehydrogenase and Phosphoglycerate kinase are coupled in vivo • Glyceraldehyde 3-phosphate dehydrogenase catalyzes an endergonic reaction while phosphoglycerate kinase catalyzes an exergonic reaction. • When these two reactions are coupled (which happens in vivo), the overall reaction is exergonic.
The formation of ATP by phosphoryl group transfer from a substrate is referred to as a substrate-level phosphorylation Substrate-level phosphorylation soluble enzymes chemical intermediates Respiration-linked phosphorylation Photophosphorylation membrane-bound enzymes transmembrane gradients of protons
2,3-Bisphosphoglycerate (BPG) • The concentration of BPG is usually low in most of the tissues except erythrocytes (up to 5 mM). • Function of BPG in erythrocytes is to regulate the affinity of hemoglobulin to O2.
10. Transfer of the phosphoryl group from phosphoenolpyruvate to ADP
Glucose + 2ATP + 2NAD+ + 4ADP + 2Pi 2 pyruvate + 2ADP + 2NADH + 2H+ + 4ATP + 2H2O Glucose + 2ADP + 2NAD+ + 2Pi 2 pyruvate + 2ATP + 2NADH + 2H+ 在有氧狀況下,產生的NADH很快就被送到mitochondria中用來合成ATP
NAD+ (nicotinamide adenine dinucleotide) is the active form of niacin
Niacin • Niacin is the common name for nicotinamide and nicotinic acid. • Nicotinic acid is the common precursor for NAD+ and NADP+ biosynthesis in cytosol.
Functions of NAD+ and NADP+ • Both NAD+ and NADP+ are coenzymes for many dehydrogenases in cytosol and mitochondria • NAD+ is involved in oxidoreduction reactions in oxidative pathways. • NADP+ is involved mostly in reductive biosynthesis.
Niacin deficiency: pellagra Weight loss, digestive disorders, dermatitis, dementia
Niacin deficiency • Because niacin is present in most of the food and NAD+ can also be produced from tryptophan (60 grams of trptophan 1 gram of NAD+), so it is not often to observe niacin deficiency. • However, niacin deficiency can still be observed in areas where maize is the main carbohydrate source because maize only contain niacytin, a bound unavailable form of niacin. Pre-treated maize with base will release the niacin from niacytin.
Niacin deficiency • Areas where sorghum is the main carbohydrate source will also observe niacin deficiency if niacin uptake is not being watched carefully. • Sorghum contains large amount of leucine, which will inhibit quinolinate phosphoribosyl transferase (QPRT), an enzyme involved in NAD+ biosynthesis from tryptophan. • Vitamin B6 deficiency can also lead to niacin deficiency because pyridoxal phosphate is a coenzyme in NAD+ biosynthesis from tryptophan.
Drug: ISONIAZID Classification: Antimycobacterial Indication: Infection with, or disease from, mycobacterium tuberculosis ISONIAZIDA Commonly Used Medicationfor HIV & AIDS Patients
Glycogen and starch are degraded by phosphorolysis • Glycogen and starch can be mobilized for use by a phosphorolytic reaction catalyzed by glycogen/starch phosphorylase. This enzyme catalyze an attack by Pi on the (a14) glycosidic linkage from the nonreducing end, generating glucose 1-phosphate and a polymer one glucose unit shorter.
Glucose 1-phosphate is converted to G-6-P by phosphoglucomutase by the same mechanism observed in phosphoglycerate mutase reaction
Digestion of dietary polysaccharides • Digestion begins in the mouth with salivary a-amylase hydrolyze (attacking by water) the internal glycosidic linkages. • Salivary a-amylase is then inactivated by gastric juice; however pancreatic a-amylase will take its place at small intestine. • The products are maltose, maltotriose, and limit dextrins (fragments of amylopectin containing a16 branch points.
Digestion of dietary disaccharides • Disaccharides must be hydrolyzed to monosaccharides before entering cells. • Dextrin + nH2O n D-glucose • Maltose + H2O 2 D-glucose • Lactose + H2O D-galactose + D-glucose • Sucrose + H2O D-fructose + D-glucose • Trehalose + H2O 2 D-glucose dextrinase maltase lactase sucrase trehalase
Lactose intolerance • Lactose intolerance is due to the disappearance after childhood of most or all of the lactase activity of the intestinal cells.
Lactose intolerance • Undigested lactose will be converted to toxic products by bacteria in large intestine, causing abdominal cramps and diarrhea.