Download

1 / 43
440 likes | 751 Vues
WNT SIGNALING IN BONE METABOLISM. J Boner Miner Metab (2009) 27: 265-271 DOI 10.1007/s00774-009-0064-8 *Takuo Kubota *Toshimi Michigami *Keiichi Ozono. GİRİŞ.

E N D
WNT SIGNALING IN BONE METABOLISM J Boner Miner Metab (2009) 27: 265-271 DOI 10.1007/s00774-009-0064-8 *Takuo Kubota *Toshimi Michigami *Keiichi Ozono
GİRİŞ • Kemik kitlesi, tepe noktasına ulaşmak için kemik yapımınınkemik yıkımını aştığı büyüme dönemi süresince artar. Yaşamın ileriki dönemlerinde kemik yıkımının yapımını aştığı süreçte osteoporoz genel bir sağlık problemi haline gelir. • Kemik kuvvetindeki kişisel farklılıklara genetik faktörler katkı sağlar.
giriş • İnsan Düşük Dansiteli Lipoprotein Reseptör Bağlantılı Protein 5 (LRP5) genindeki fonksiyon kaybı ve kazancının sırasıyla Osteoporosis-pseudoglioma sendrom (OPPG) ve Yüksek kemik kitlesi (HBM) fenotipi ile bağlantılı olduğu gösterilmiştir. Bu bulgular Wnt sinyalinin kemik metabolizmasındaki rolüne ışık tutar. Çünkü LRP5 Wnt sinyalinin ko-reseptörüdür.
Wnt Sinyali Salgılanmış Sistein-zengin glikoproteinler olan Wnt proteinleri en az üç farklı yolla aktive olur: 1. Doğal (β-catenin bağımlı) 2. Ca*² 3. Planar polarite Bu üç yoldan Doğal yol en iyi anlaşılmış olanıdır.
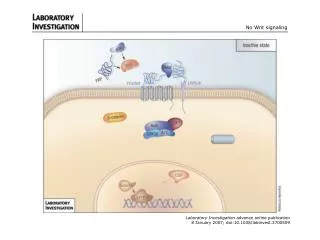
Doğal Wnt Sinyal Yolağının Şeması Dickkopfs (Dkks), Secreted frizzled-related proteins (Sfrps) ve Sclerostin (SOST geni ürünü) salgılanmış Wnt inhibitörleridir.
Doğal Wnt Sinyal Yolağının Şeması Wnt sinyalinin eksikliğinde β-catenin, glycogen sentaz kinaz 3β (GSK3β) ile fosforile edilerek proteozomal degradasyonu başlatır.
Doğal Wnt Sinyal Yolağının Şeması Hücre yüzeyinde Wnt sinyali, Frizzled ve LRP5/6 ile üçlü kompleks şeklinde bağlandığında GSK3β inhibe, β-catenin stabilize olur.
Doğal Wnt Sinyal Yolağının Şeması Sonrasında β-catenin çekirdekte birikir ve lenfoid geliştirici faktör (LEF)/ T-hücre faktörü (TCF)-aracılı gen transkripsyonunu aktive eder.
Wnt sinyali • Wnt sinyali için diğer bir koreseptör olan LRP5 de LRP6 gibi tek geçişli transmembran proteinidir. • Bunlar hücre dışında 4 adet YWTD β propeller (pervane) alandan oluşur. Bu yapılar Wnt sinyalinde ko-reseptör olarak işlev görür. • Endoplazmik Retikulumdaki bir şaperon proteini olan Mesoderm Development (Mesd) in hücre yüzeyindeki LRP5/6 trafiği için gerekli olduğu rapor edilmiştir. • Doğal Wnt sinyali sadece embriyo gelişiminde değil, aynı zamanda yetişkin dokuların korunması ve hastalıkların artışı bakımından da önemlidir.
LRP5 • LRP5 Geninin fonksiyon kaybına neden olan mutasyonlar nadir görülen OR bir bozukluk olan, erken osteoporoz ve körlük ile karakterize OPPG ye neden olur. Bu hastalarda kemik kitlesi azalması ve iskelet frajilitesi görülür. • OPPG ye zıt olarak LRP5 genindeki fonksiyon artış mutasyonları OD olan HBM fenotipi ile bağlantılıdır.
Lrp5 • Yakın zamanda yayınlanan büyük bir genom çalışması ve geniş çaplı analizi, bize LRP5 varyantlarının kemik mineral dansitesi ve kırık riski ile bağlantılı olduğu göstermişti. LRP5 eksikliği mutasyonları olan farelerde insan kemik fenotipleri kopyalandı ve HBM fenotipinde LRP5^G17IV olarak korunduğu ifade edildi. • LRP5¯ ́¯ olan farelerde düşük kemik kitlesi ve azalmış osteoblast proliferasyonu ile fonksiyonu gösterildi.
Lrp5 • Aşırı üretilmiş LRP5^G17IV proteini olan fare osteoblastlarında artmış fonksiyon ve azalmış apopitozise bağlı olarak kemik hacmi ve kuvveti de artmıştı. • Mutant tip LRP5 aşırı üretimi kemik hacminin aşırı derecede artmasına olanak verir. • İnsan ve fareler üzerinde yapılan bu çalışmalar LRP5 in osteoblast proliferasyonu, differansiasyonu, fonksiyonu ve apopitozisi üzerinden düzenleme yaparak osteogenez üzerinde kritik bir role sahip olduğunu gösterdi.
Lrp5 • LRP5¯ ́¯ olan farelerde intermittan PTH ya yanıt olarak kemik formasyonu artar. Böylelikle LRP5 in PTH ya bağlı kemik metabolizmasında gerekli olmadığı görüldü. • PTH/Protein Kinaz A sinyalinin GSK3β yı inaktive ederek Doğal Wnt yolağını aktive ettiğigösterilmişti. İşte bu yol PTH nın anabolik yolaklarından biridir.
Lrp5 • LRP5¯ ́¯ farelerde ulna’nın mekanik yüklenmeye bağlı osteogenik cevabı mutant tip farelerin osteogenik cevabına göre azalmış bulundu. Doğal Wnt sinyalindeki hedef genler canlıda tibia yüklenmesinden sonra geliştirildi. • Bu sonuçlar kemikte mekanik yüklenmeye LRP5 in aracılık ettiğini göstermiştir.
Lrp5 • LRP5 hipermorfik ve hipomorfik mutant proteinleri fonksiyonel olarak incelendi. Hipermorfik LRP5^G17IV HBM mutasyonunun plazma membranına hedeflenmenin azalmasına ve Mesd ile etkileşimininbozulmasına neden olduğu bildirildi. • Ne var ki; diğer gruplar, ilk YWTD alanındakümelenme mutasyonunauğrayan LRP5 HBM proteinlerinin plazma membranına ulaşımını sağladığını, Dkk1 ve sclerostin yoluyla inhibe olarak Wnt sinyali gelişimine affinitesinin azaldığını göstermiştir.
Lrp5 • Diğer yandan OPPG ye sokulan LRP5 in bazı missense mutasyonlarındaki hipomorfizmin, Doğal Wnt sinyal transdüksiyonu azalmasını ve hücre yüzey bozulmasını başlattığı, her ne kadar tamamen aydınlatılmasa da, rapor edildi.
LRP6 • LRP6 eksik olan mutant farelerdeki komposit defektlerin Wnt1, Wnt3a ve Wnt7a’yı içeren çeşitli Wnt genlerindeki mutasyonlardan kaynaklandığı ve perinatal dönemde ölüme sebep olduğu gösterilmişti. • LRP6 eksik allellerdeki heterozigositenin kemik mineral yoğunluğunu LRP5 eksik farelere oranla daha fazla azalttığı rapor edildi.
Lrp6 • Önceden belirlediğimiz bir nokta mutasyonu olan ringelschwanz (rs), LRP6 geninde spontan missense mutasyonu olan farelerde somitogenes defektine, ossifikasyon merkezlerinin ortaya çıkmasında gecikmeye ve yetişkin kemik kitlesinin azalmasına neden oldu. • LRP6^rs/rs farelerde azalmış trabeküler kemik kitlesi ile birlikte genişlemiş erode yüzey gösterildi.
Lrp6 • LRP6^rs/rs farelerde üriner deoksipridinolin atılımı daha yüksek iken serum osteokalsin seviyeleri mutant tip ikiz eşleri ile karşılaştırılabilir seviyedeydi. • LRP6^rs/rs osteoblastlarda belirgin bir proliferasyon, mineralizason ve diferansiyasyon defekti gösterilemedi. • Reseptör aktivatör nükleer faktör-κ B ligand (Rankl) ifadesi in vivo ve in vitro incelemelerdeLRP6^rs/rs osteoblastlarda geliştirilir ve osteoklastogenez ile kemik yıkım aktivitesi hızlandırılır.
Lrp6 • LRP6 aşırı üretimi aynı zamanda Rankl ürünlerini baskılarken, LRP6^rs mutant proteini baskılamaz. Benzer şekilde 7-kb RANKL promoter aktivitesi β-catenin aşırı üretimi ile azaltılır. • Bununla birlikte; LRP6 aracılı sinyal, postnatal kemik kitlesini kontrol ederken bir parça da kemik rezorpsiyonunu ayarlar. • İnsanlarda yapılan bir çalışmada LRP6 geni mutasyonu sonucu osteoporoz ile erken koroner hastalık, hiperlipidemi, hipertansiyon ve diabet birlikteliği görüldü.
Lrp6 • LRP6^rs mutantları üzerinde yapılan fonksiyonel analizler LRP6 için bir şaperon olan Mesd etkileşiminin azalması ile, Wnt/β sinyalinin hasarını başlattığını ve plazma membran hedeflenmesini azalttığını açığa çıkardı. • Mesd etkileşimi LRP6 fonksiyonu için olduğu gibi LRP5 fonksiyonu için de kritik bir öneme sahiptir. Mesd, LRP5/6’nın hipomorfik mutasyonlarında terapotik hedef molekül olabilir.
WNT • Wnt metabolizmasında görevi tamamen açıklanamamış 19 üye vardır. Osteoblastlar, Kondrositler, Myositler ve Adipositler mezenşimal kök hücrelerden köken alır. Adipogenezis, diğer hücrelere dönüşüm sinyali ile uyarılmayan mezenşimal kök hücrelerinde gelişir. • Wnt1 ve Wnt10b periadipositlerin adipogenezisini inhibe eder.
Wnt • Wnt7b, Wnt10b ve Wnt5a içeren bazı Wnt’ler farelerde kemik hemostazında fonksiyon gösterir. Wnt7b, osteoblastlarda kodlanır ve osteoblast diferansiyasyonu ile ifade edilir. • Wnt7b’nin kaldırılması, Dermo-Cre kullanan fare iskelet progenitör hücrelerinde kondrogenez ve osteoblastogenez defekti ile sonuçlanır. Wnt7b’nin Gα q11-protein kinaz Cδ yolağıyla (Doğal Wnt sinyal yolağı aracılığıyla değil) osteoblast diferansiyasyonuna neden olduğu gösterildi.
Wnt • Transgenik Wnt10b fareler Adiposit-spesifik FABP4 promoterini kullanarak kemik kitlesini yükseltir ve yağ dokusunu azaltır. • Wnt10b, osteoblastik proliferasonuna osteoblastogenik transkripsiyon faktörleri olan Runx2 ve Osterix’i indükleyip; adipogenik transkripsiyon faktörleri olan CCAAT/ enhancer binding protein α (C/EBPα) ve Peroxisome proliferator-activated receptor γ ( PPARγ) yı baskılayarak etki eder. • Osterix, doğal wnt sinyalini inhibe eder. Farelerde osteokalsin promoterini kullanarak Wnt10b aşırı üretimi yapılması osteoblastogenezi stimüle ederek kemik kitlesini arttırır.
Wnt • Wnt10b¯ ́¯ farelerde kemik formasyonu azalmasıyla trabeküler kemik azalır. Bu sonuçlar Wnt7b ve Wnt10b’nin kemik formasyonunun endojen regulatörleri olduğunu gösterir. • Wnt5a* ́¯ farelerde kemik iliği mezenkimal progenitörlerinde gelişmiş adipogenez ve azalmış osteoblastogeneze bağlı olarak kemik kitlesi azalır. Wnt5a, kalsiyum/calmodulin bağımlı protein kinaz ІІ yoluyla, PPARγ transaktivasyonunu histon metiltransferaz aracılığı ile kromatin inaktivasyonu yaparakbaskılar.
SFRP • Sfrp’ler de Wnt’ler ile Frizzled reseptörler arasındaki etkileşimi önleyen Wnt antagonistleridir. Sfrp1¯ ́¯ yetişkin fareler trabeküler kemik artışı gösterir. Sfrp1 delesyonu in vivo çalışmalarda osteoblast apopitozisini inhibe ederken, in vitro düzeyde osteoklastogenezi arttırır.
DKK • Dkk’lar salgılanmış glikoproteinlerdir. Dkk1, Dkk2 ve Dkk4; LRP5 ve LRP6 ile etkileşimdedir. • Wnt/β-catenin sinyalini inhibe eder. Dkk1¯ ́¯ fareler hatalı baş indüksiyonu nedeniyle ölür. • Dkk1* ́¯ fareler kemik rezorpsiyonunu değiştirmeden kemik formasyon artışı ile HBM fenotipi gösterir. • Fare osteoblastlarında aşırı üretilmiş Dkk1, azalmış osteoblast sayısı ve kemik formasyonuna bağlı osteopeni gelişimine neden olur.
Dkk • Dkk1 antisense oligonükleotid tedavisi, overyektominin mineral dansitesi üzerindeki baskılayıcı etkisini ve femurlardaki yükün tepe noktasına ulaşmasını önler. Osteoblast sayısını arttırır. Rankl üretimini baskılayarak osteoklast diferansiyasyonunu bozar. • Dkk1, kemik formasyonuna negatif etkide bulunur ve osteoklastogeneze indirekt yolla etki eder.
Dkk • Beklenmedik şekilde, Dkk2¯ ́¯ fareler artmış osteoid yapı ve azalmış mineralizasyon nedeniyle osteopeniktir. Bunlar; Rankl üretiminin arttırılması ile gelişen osteoklastogenez nedeniyledir. • Böylece Dkk2 nin kemik formasyon ve rezorpsiyonuna etkili olduğu görülür. Doku analizleri ve mutagenez çalışmaları fare Dkk2’lerinin C-terminal sistein zengin alanlarının üçüncü LRP5/6 β propeller alanları ile bağlantılı olduğunu göstermiştir.
SCLEROSTİN • Sclerostin, SOST geni tarafından kodlanıp salgılanan Wnt antagonistidir. Birçok çalışmada sclerostinin LRP5 ve LRP6 ya bağlanıp Wnt/β-catenin sinyalini ve kemik morfogenetik proteini (BMP) yolağını inhibe ettiği gösterilmiştir. • LRP5^G17IV mutasyonu, sclerostin-LRP5 bağlantısını iptal edip sclerostin ile inhibisyona direnç sağlayarak HBM fenotipine yatkınlık gösterir.
Sclerostin • İnsanda fonksiyon kaybı mutasyonları ve SOST geni ürünlerinin azalması sırasıyla Sclerosteozis ve Von Buchem hastalığı ile birlikte anılır. Farelerde SOST geni aşırı üretimi kemik kitlesi ve formasyonunda sırasıyla artış ve azalmaya neden olur. • Sclerostinin osteositlerde açıklanamayan bir şekilde kemik remodelingi ve mekanik yük algısı kaynaşmasını sağladığı öne sürülmüştür. Böylece, Sclerosin’inumut vaat eden terapotik hedef molekülolduğu söylenebilir.
Sclerostin • Sclerostin monoklonal antibody’ler postmenopozal kadınlarda kemik formasyon işaretçilerini geliştirir ve arttırır. • Mekanik stimülasyon ve PTH tedavisi sclerostin üretimini azaltıp endojen wnt inhibisyonunu hafifleterek kemik formasyonunu arttırabilir. • Transgenik fare osteositlerinde aktif PTH reseptörü kemik kitlesi ve remodelingi arttırır. • LRP5 delesyonu HBM fenotipini azaltır. Fakat transgen nedenli kemik remodelingini arttırmaz. Osteositlerde PTH reseptör sinyali LRP5 bağımlı ve bağımsız mekanizmalarla kemik metabolizmasına etki eder.
GSK3β • GSK3β, β-catenin ile birlikte birçok substratı hedefleyen bir serin/treonin kinazdır. • Bir GSK3β inhibitörü olan Lityum, farelerde kemik kitlesi ve formasyonunu arttırır. • Şaşırtıcı bir şekilde bir vaka kontrol çalışmasında Lityum tedavisi insanlarda azalmış kemik kırığı riski ile birlikte anılmıştır. • GSK3β* ́¯ farelerde gelişen Runx2 transkripsiyonel aktivitesine bağlı olarak kemik formasyon artışı gösterilmiştir.
β-CATENİN • β-Catenin, osteoblast gelişiminin çeşitli evrelerinde farklı roller oynar. • Dermo1-Cre fareler ve Prx1-Cre farelerin kondrosit ve osteoblastlarının mezenkimal prekürsörlerindeki β-catenin delesyonu osteoblast diferansiyasyonunu bloke eder. • İntramembranöz ve enkondral kemikleşme boyunca kondrositlerde ektopik kemik formasyonuna neden olur.
β-catenin • Böylece β-Catenin’in osteoblast formasyonunun erken evrelerinde gerekli olduğu görülmüştür. Olgun osteoblastlarda β-Catenin başka önemli fonksiyonlarda da rol oynar. • α1(1)collage-Cre fare osteoblastlarında β-Catenin inaktivasyonu kemik rezorpsiyonunu arttırıp osteoproteogerin (opg) üretimini geliştirerek kemik kitlesinde azalma başlatır.
β-catenin • Ek olarak Osteocalsin-Cre farelerde β-Catenin kaybı osteoklast artışı ile şiddetli osteopeniye neden olmuştur. Doku incelemelerinde β-Catenin eksik osteoblastlarda artmış Rankl üretimi ve azalmış Opg üretimi görülmüştür. • Bu bulgular, olgun osteoblastlarda β-Catenin’in osteoklastogenez ve osteoklast fonksiyonunu ayarladığını gösterir.
TCF • Tcf’ler, Wnt/β-Catenin sinyaline aracılık eden transkripsiyon faktörleridir. • Tcf1¯ ́¯ farelerde düşük kemik kitlesi görülür. Bu mutant farelerde osteoblast sayısı ve fonksiyonu değişmezken osteoklast sayısı ve opg üretimi artmıştır. • Bu bulgular diğer spesifik β-catenin eksik fare osteoblastlarındaki bulgularla benzerdir. • Oysa bir başka çalışmada; doku Runx2 geninin TCF bağlanma alanından doğal Wnt sinyal yolağını direkt olarak hedef aldığı gösterilmiştir.
ÖZET Canlılar üzerinde yapılan çalışmalardaki çeşitlilikle LRP5 genindeki mutasyonların bulunması Wnt sinyalinin kemik metabolizmasındaki rolünü anlamamızı sağlamıştır. Osteoblastlardaki Lrp6-aracılı sinyal osteoklastogenezi düzenleyebilir. Ne var ki; Osteoklastlardaki Doğal Wnt sinyalinin rolü tam olarak açıklanamamıştır. Doğal olmayan Wnt sinyalinin kemik metabolizmasındaki rolünü anlamada da bilgilerimiz kifayetsizdir.
Doğal Wnt Sinyalinin Osteoblast Diferansiyasyonundaki Rolü Doğal Wnt sinyali mezenkimal progenitör hücrelerinin osteoblastlara dönüşümünü arttırırken kondrosit ve adiposit diferansiyasyonunu inhibe eder.
Doğal Wnt Sinyalinin Osteoblast Diferansiyasyonundaki Rolü Osteoblastlarda, Wnt yolağı OPG/ RANKL oranını arttırarak apopitozis ve osteoklastogenezisi bloke ederken proliferasyon ve mineralizasyonu arttırır.
Doğal Wnt Sinyalinin Osteoblast Diferansiyasyonundaki Rolü Sclerostin, osteositlerde üretilen, osteoblast proliferasyonunu ve fonksiyonunu baskılayan bir doğal wnt sinyali inhibitörüdür.
Özet Farmakolojik açıdan, diğer dokularda tumörigenisite ve doku toksisitesi önemli olmasına rağmen Wnt sinyal yolağındabirçok basamak terapotik hedef olarak dikkate alınabilir. Plasma membranındaki LRP5/6 miktarını arttırmada Mesd hedef moleküllerden biri olabilir. Sclerostin, osteositlerde bilinmeyen bir şekilde kodlandığı için, sclerostin oluşumunun önlenmesi en umut verici tedavi yöntemidir.