Download
1 / 52
611 likes | 1.46k Vues
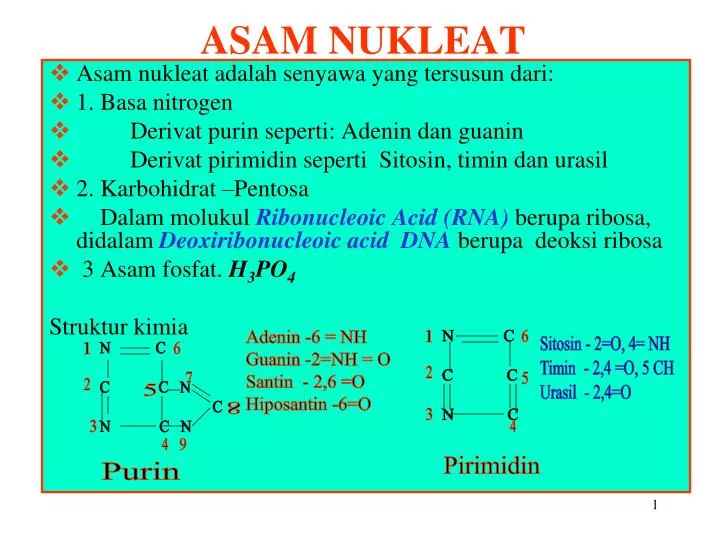
ASAM NUKLEAT. Asam nukleat adalah senyawa yang tersusun dari: 1. Basa nitrogen Derivat purin seperti: Adenin dan guanin Derivat pirimidin seperti Sitosin, timin dan urasil 2. Karbohidrat –Pentosa

E N D
ASAM NUKLEAT • Asam nukleat adalah senyawa yang tersusun dari: • 1. Basa nitrogen • Derivat purin seperti: Adenin dan guanin • Derivat pirimidin seperti Sitosin, timin dan urasil • 2. Karbohidrat –Pentosa • Dalam molukul Ribonucleoic Acid (RNA) berupa ribosa, didalam Deoxiribonucleoic acid DNAberupa deoksi ribosa • 3 Asam fosfat. H3PO4 Struktur kimia Adenin -6 = NH Guanin -2=NH = O Santin - 2,6 =O Hiposantin -6=O 1 N C C C N C 6 Sitosin - 2=O, 4= NH Timin - 2,4 =O, 5 CH Urasil - 2,4=O 1 N C C C N C N C N 6 2 7 5 2 5 8 3 3 4 4 9 Pirimidin Purin
Berbagai basa turunan purin dan pirimidin 1 N CH C H C N CH 6 6 N CH C C N CNH 1 NH N C C C N C N C N 2 Adenin 5 7 H 2 5 O= H H 2 H 8 H H 3 4 3 H 4 9 Sitosin Pirimidin H N C C HC N CH N HC N H Guanin N CH O=C CH N C =O N CH O=C C -OH N C =O HN= H H H H Urasil Timin H = O N C C HC N CH N HC N H N C C HC N CH N HC N H H = O H O = H Hiposantin Santin
Turunan Ribose O HO- P-O- CH O HO-CH HO-CH 2 2 2 O O O H OH OH H • Contoh OH H H OH H OH H OH H OH H OH H H OH O= P- O Ribose monofosfat D-Ribose Deoksi D-ribose O O= P- O O O- P-O- CH O O HO- P- O O O- P-O- CH O 2 O O Adenosin H OH H 2 D-ribose difosfat H OH H OH H OH H OH Ribose nukleotida trifosfat
O O OH OH H HO- P - O -CH- C - C - C - C OH H H H O N C C C N C N C N • Asam nukleat • Adenilat ( Adenosin monofosfat) • Nukleosid adalah nukleotida yang dihilangkan asam fosfatnya. Sehingga tata nama: • Asam adenilat • Asam uridinilat. Uridinilat H 2 O O N C C C N C H O OH OH O C - C - C - C - CH - O - P - OH H H H H OH O H 2 Nukleosid Nukleotid 5
Fungsi nukleosid dan nukleotida A.Sebagai nukleosid dan nukletid 1.Penyusun ATP (adenosin Trifosfat), dan ADP atau Adenosin Difosfat. GTP = Guanosin Trifosfat, GDP = Guanosin Difosfat 2. S-Adenosil metionin (metionin yang aktif), PADS = 3-fosfoadenosin-5’-fosfosulfat = Sufat aktif. B. Sebagai senyawa yang bekerja sebagai koenzim: 1. Kodehidrogenase FAP(Flavine Adenosin Dinukleotida) NAD = Nikotinamid Adenosin Dinukleotida,: dan Nikotinamid Adenin Dinukleotid =NADP. 2. Koenzim A-SH (Pantetein berikatan dengan Adenosin 3 fosfat. AMP siklik dari ATP yang dikatalisis oleh adenilat siklase, berperan dalam glukogenolisis dan glukogenesi oleh hormon efinefrin
Komponen penyususn Struktur kimia purin 7 N C N 6 5 N H N-C N 1 ADENOSIN Biosintesis purin Penelitiaanya menggunakan radio aktif. Dengan tahapan: 1. Atom C no 6, intipurin berasal dari nolekul CO2 dari udara. 2. Atom (N=1), inti purin berasal nitrogen asam aspartat. 3. Atom C no.2, dan atom C no.8 adalah produk dari reaksi transformilasi yang berasal dari senyawa donor gugus for- mil, yang mengkaitnya koen- zim FH4 (tetrahidrofolat). 8 4 2 3 2 9 O HC CH HC CH CH 2 O O=P - O Pentosa Ribosa O HMP-Siklik As. Fofat
4. Atom N (3), dan N (9), dari nitrogen gugus amida molekul glutamin. 5. Atom C (4) dan C (5), dan N (7), merupakan molekul glisin seluruhnya. Tahapan reaksi sintesis purin sebagai berikut: 1. Pembentukan PRPP (5-fosfo Ribosil Piro fosfat), dengan bantuan ATP, dan ion Mg++ sebagai aktivator 2. Kemudian pembentukan 5’fosforibosilamin dari reaksi PRPP, dengan glutamin akan menghasilkan asam glutamat dan Ppi. 3. Berikutnya pembentukan Glisin Amid Ribosil-5P (GAR), hasilnya adalah Ribosilamin 5P, dengan glisin. Dalam reaksi ini dikatalisis oleh ATP dan aktivator Mg++., dan dibantu oleh enzim GAR-sintetase GAR menenpati posisi 4,5,7 dan 9 dari inti purin 4. GAR melakukan formilasi dikatalisis oleh enzim transformilase, dengan koenzim FH4. Dan senyawa donor gugus formil terjadi formil glisinamidinrebosil 5P.
Atom C dari formil menduduki C-8 dari purin • 6. Formilamid-ribosil-5P mengalami aminasi pada ato C-4, dengan senyawa donor amin (glutamin), terbentuklah formil-glisinamid-ribosil-5P. Aminasi ini membentuk N (3) pada inti purin. • 6. Selanjutnya terjadi siklisasi, dengan membentuk amino-imidazol-ribosil-5P. Senyawa ini bereaksi dengan CO2 udara dengan biotin sebagai koenzim dan atom C menenpati C(6) pada intipurin. Dengan asam aspartat membentuk senyawa 5-amino-4-imidazol-N-suksinil-karboksamid-ribosil-5P. • 7. Senyawa 5-amino-4-imidazol-N-suksinil-karboksamid-ribosil-5P. Diubah menjadi 5-amino-4-imidazol-karboksamid -ribosil-5P, dan asam fumarat.dengan dikatalisisi oleh enzim adenosilsuksinase • 8. Senyawa5-amino-4-imidazol-karboksamid-ribosil-5P, mengalami formilasi, dengan pendonor FH4-dan terbentuk senyawa 5-formamido-4-imidazol-karboksamid-ribosil-5P. Gugus formil menempati C (8), inti purin.
9. Tahap akir terjadi reaksi siklisasi cincin ke dua, dan terbentuk derivat purin pertamma yang dinamaan inosin monofosfat (IMP= asam inosinat), dinamakan juga hiposantin atau 6-oksi purin yang kemudian dapat diubah adenosin monofosfat (AMP atau GMP = Guanosin monofosfat. PRPP( 5-fosforibosilpirofosfat) HCO P-O-P- (OH) HCOH HCOH HC- H C-O-P HC-NH HCOH HCOH HC- H C-O-P 2 2 O HCOH HCOH HCOH HC- H C-O-P ATP Mg AMP Gltamat + PPi O O Gltamin 2+ H NOC(CH ) CH(NH )-COOH 2 2 2 2 2 2 5-Fosforibosilamin 2 2 H C NH O=C NH R-5p Glisin Ribose-5P GAR(Glisinamid-ribosil-5P)
5-amino-4-imidazol-N-suksinil- karboksamid-ribosil-5P 5 7 ATP H C NH CHO O=C NH R-5P 5 7 2 8 H C NH CHO H NC NH R-5P Siklisasi H C N CH H NC N R-5P Glutamin 2 8 2 9 4 2+ 9 Mg 4 2+ Mg 2 ATP 2 Glisin ribose 6-fosfat ATP 2+ Biotin Mg As. aspartat 6 HOOC H C HC-NH-C=O HOOC CO 1 2 H N-C =O 7 2 2 5 C N CH H NC N R-5P 8 HCHO.transformilasi C N CH H NC N R-5P 3 9 HOOC HC CH HOOC 4 2 2 Fumarat 6 1 5-amino-4-imida zol-arboksamid -ribosil-5P H N C=O O=C C N CH H N C N R-5P 6 1 2 N C-OH H C C N CH N C N R-5P 7 5 2 7 8 NADPH 2 5 8 9 2 4 3 2 9 4 3 Inosin monofosfat
HOOC-CH CHCOOH 2 ATP N C-NH H C C N CH N C N R-5P Aspartat 2 H O 2 NAD • Inosin monofosfat Adenilsuksinat N C-OH HOC C N CH N C N R-5P Santinmonofosfat) N C-NH H C C N CH N C N R-5P HOOC-CH= CHCOOH 2 Fumarat NAD 2 N C-OH H NC C N CH N C N R-5P N C-OH O=C C N CH N C N R-5P +NH 3 Glutamin 2 ATP AMP (adeninmonofosfat) GMP (Guaninmonofosfat)
Pengaturan Biosintesis Ribonukleotida • Lintasan sintesis purin dan pirimidin de novo terutama diatur oleh konsentrasi produk-produknya. Bentuk pengaturan ini menjamin suplai nukleotida yang adekuat dan mencegah produksi yang berlebihan. • Kedua lintasan juga dipengaruhi oleh konsentrasi PRPP. Besarnya mol PRPP terutama dipengaruhi oleh kecepatan sintesisnya melalui ribosa-fosfat pirofosfo kinase dan kecepatan pengunaannya oleh lintasan penyelamatan (Iihat bawah). • Pada individu normal, ribosa-fosfat pirofosfokinase secara umpan-balik dihambat oleh nukleotida purin dan pirimidin (slide 16). Individu dengan bentuk ribosa-fosfat pirofosfokinase yang abnormal akan superaktif atau resisten terhadap penghambatan umpan-balik.
Enzim pertama dan menghambat-kecepatan lintasan purin, glutamin PRPP amidotransferase, dihambat secara umpan-balik oleh sejurnlah nukleotida purin, yaitu IMP, ADP, GMP, dan GDP (slide 16) • Akan tetapi, bila konsentrasi PRPP intraselular meningkat melampaui kadar normal, glutarnin PRPP amidotransferase terlepas dari penghambatan umpan-balik, dan pengaturan lintasan purin terganggu. • Jadi, individu dengan bentuk ribosa-fosfat pirofosfokinase yang superaktif atau resisten terhadap umpan-balik membentuk nukleotida purin yang berlebihan. • Pengatur tambahan pada lintasanpurin mengatur pembentukan AMP dan GMP dari IMP. Nukleotida adenin dan guanin masing-masing menghambat pro duksinya sendiri. (Alosterik)
Sintesis piridin terlihat bahwa Enzim pertama lintasan pirimidin, karbamiol fosfat sintase II, dihambat secara umpan-balik oleh UTP dan dirangsang oleh ATP. • Jadi, bila kadar ATP meningkat,mereka merangsang peningkatan pernbentukan pirimidin. CTP sintase dihambat secara umpan-balik oleh produknya sendiri, yaitu CTP. • Berbeda dengan biosintesis Deoksiribonukleotida yang diperlukan untuk sintesis DNA dibentuk dari ribo nukleotida oleh reduksi cincin pentosa pada posisi 2'. • Salah satu enzim, ribonukleotida reduktase, yang mengkatalisis perubahan masing-masing ribonukleotida difosfat menjadi deoksiribonukleotida difosfat yang sesuai. NADPH menyumbangkan ekuivalen pereduksi yang digunakan pada reaksi ini melalui perantara yang tidak diketahui. (Slide 19)
Enzim redukse Ribonukleotida Sintesis deoksiribonukleotida dikatalisis oleh ribonukleo tida reduktase. Ekuivalen pereduksi yang digunakan pada reaksi ini disediakan oleh NADPH dan dipindahkan ke ribonu kleotida melalui karier yang tidak diketahui
Aktivitas ribonukleotida reduktase diatur sehingga men jamin bahwa sintesis deoksiribonukleotida seimbang, yaitu, tidak satu pun deoksiribonukleotida disintesis dalam jumlah yang tidak sesuai dengan lainnya (yang diperlukan). • Jenis pengaturan ini penting karena ketidak seimbangan pada pool deoksiribonukleotida akan menurunkan ketepatan sintesis DNA, dan akibatnya akan meningkatkan kecepatan mutasi. • Ribonukleotida reduktase secara alosterik diaturoleh ATP, dTTP, dGTP, dan dATP ATP meningkatkan kecepatan enzim yang mengubah sitosin difosfat (CDP) dan uiridin difosfat (UDP) menjadi deoksiribonu kleotida yang sesuai. • DeoksiUDP melalui beberapa langkah, diubah menjadi dTTP/.
Yang terakhir ini menghambat ribonukleotida reduktase yang sesuai dengan CDP dan UDP tetapi mengaktifkannya ribonukleotida reduktase yang sesuai dengan guanosin difosfat (GDP) dan adenosin difosfat (ADP). • Jadi, pirimidin deoksiribonukleotida secara umpan-balik menghambat pembentukannya sendiri dan merangsang sintesis purin. • Hal yang sarna, dGTP menghambat produksinya sendiri' dan pirimidin merangsang pembentukan dATP. Akhirnya, dATP menghambat enzim yang sesuai dengan semua (keempat) substrat ribonukleotida, jadi memintas (shunt off) sintesis deoksiribonukleotida. • Sebagai akibat lintasan penyelamatan yang overaktif, kelebihan dATP atau dGTP akan tertimbun. Penimbunan ini mencegah sintesis pirimidin deoksiribonukleotida, dengan akibat sintesis DNA dihambat.
Pengaturan biosintesis purin Dalam sintesis purin setelah terjadinya PRPP, kemudian menjadi IMP diperlukan (1)asam glutamat, (2) glisisn, (3) meteniltetrahidrfolat dan (4) adalah asam aspartat maupun CO2 sebagai prazat. Dalam sintesis itu memerlukan tenaga sangat besar, sehingga pengaturan sintesis purin kecepatannya perlu ada penga turan. Pengatur tunggal yang penting adalah PPR-ribosa-P, yang bergantung pada kecepatan sintesis, dan penggunaan. Kecepatan penggunaan PPRibose-P, sangat dipengaruhi oleh lintas penyelamatan fosforilasi hiposantin dan guanin menjadi ribonukletida, sehingga masing-masing dapat sebagai umpan balik penghambatan alosterik dalam sintesis PPR menjadi IMP yang bagannya sebagai berikut,
Bagan pengaturan kecepatan sintesiss purin PP-Ribosa-P sintetase R5-P + ATP. PP-Ribosa -P 5-Fosforibosilamin GAR (Glisisnamid ribosil -5P) 5-Amino-4-imidazol-N-suksinil-karboksamid ribosil-5P IMP AMP GMP ADP GDP ATP GTP
Nukleotida yang mengandung timin berasal dari reaksi deoksiuridin monofosfat (dUMP) mengalami metilasi membentuk deoksitimidin monofosfat (dTMP) (slide 13). • Timidilat sintase mengkatalisis pemindahangugus metilen dari N5 ,N10 -metilen-H4folat ke dUMP dan serentak mereduksi gugus yang dipindahkan oleh kofaktor folat. • Reaksi ini jarang terjadi pada reaksi yang memerIukan folat, karena dalam proses ini, kofaktor dioksidasi menjadi H2 folat. Sebelum H2 folat dapat digunakan kembali pada pemindahan satukarbon, ia harus direduksi oleh dihidrofolat reduktase.
Reaksi katalitis penyelamatan purin Reaksi katalitis purin 25
Lintasan penyelamatan purin • Nukleotida dihidrolisis oleh 5’nukleotidase akan dilepaskan Pi (pirofosfat) menjadi gunaosin, atau adenosin. • Adenosin mengalami adenosin deaminasi oleh enzim adenin deaminase inosisn atau deoksiinosin. • Bila kedua senyawa tersebut dipengaruhi oleh enzim purinnukleosida fosforilase dapat melepaskan ribose 5’-fosfat dan menjadi hiposantin atau guanosin. • Hasil ini bila dipengaruhi oleh hiposntin-guanin fosfori bosiltransferase dapat menjadi IMP, GMP maupun AMP Hasil penelitian difisiensi genitik bila tidak ada enzim adenosin deaminase atau enzim purin nukleosida fosforilase akan menyebabkan patologis. Kekebalan infeksi tak ada pada individu yang kekerangan enzim tersebut.
O NH N O NH N N N H Dalam bagan terlihat IMP yang terbentuk dapat diubah menjadi GMP maupun AMP. IMP sendiri dapat diubah oleh enzim adenosin kinase sehingga membentuk dAMP. (deoksiadenosin monofosfat). Dan terfosforilasi lagi menjadi dATP. Contoh metabolisme lain: N N H Guanin Hiposantin NH 2 O 2 O NH N H Ksantin oksidase Guanase N N H Ksantin NH = O 3 Alantoin Ksantin oksidase O 2 N N H O NH N O NH N H H N O N H H N C O N H 2 H N N H O Urikinase O = = O Asam urat [O] + H O CO Alupurinol
Keterangan • Bila alantoin yang mudah larut ini dapat terjadi maka tak akan ada penyakit asam urat. Karena itu, manusia, burung reptil dan dan amfibi produk akhir dari katabolisme purin adalah asam urat dan guanin. • Disosiasi asam urat sangat tergantung pH tubuh yang umumnya dibawah 7,0 (5,8) maka asam urat akan tetap sebagai asam. Urin dengan pH 5,0 hanya dapat melarutkan 15 mg/ dl. (desi liter). • Sedangkan pH urin 5,8, maka ditemukana sebagai natrium asam urat didaerah proksimal. Untuk memaksimalkan pengeluaran asam urat dengan mengakaliskan urin. • Beberapa kelainan katabolisme purin telah dicatat, salah satunya adalah hiperurisemia, bila pengeluaran asam urat total dalam 24 jam lebih dari 600 mg/24 jam. • Penderita dapat ditemukan antara 2000 s/d 4000 mg/24 jam dan bahkan yang berat sampai 30.000mg/24 jam.
Katabolisme ksantin menjadi asam urat, diekskresi lewat urin, bila berlebih dalam plasma disebut hiperurikemia, dan dapat mengendap dalam jaringan sebagai Na urat. • Pengambilan asam urat oleh makropag menimbulkan sindrom gout (peradangan) yang kuat. Asam urat dapat mengendap dijaringan sendi. Hal ini akibat katabolisme ksantin yang terlalu cepat. • Defisiensi ribose-fosfatpirofosfokina sedan HGPRT ase akan menyebabkan PRPP berlebihan akibatnya terjadi peningkatan glutamin PRPP amididotransferase, berle bihan dan tak terhambat, sehingga purin over produksi. • Individu yang mengalami difisiensi HGPRT ase parsialakan menderita gout, tetapi kalau mengalami defisiensi total akan menyebabkan sindroma Lesch-Nyhan. Dapat mengalami retardasi mental dan dapat merusak diri sendiri. • Aluperinol mempunyai struktur yang mirip asam urat dapat digunakan untuk mengobati hiperurisemia, karena dapat menghambat ksantin oksedase, sehingga tak terjadi asam urat, tetapi berhenti di ksantin dan hiposantin.
Biosintesis Pirimidin Seperti halnya purin, mempunyai baha asal seperti PROO, glutamin, CO2, asam aspartat, koezim tetrahidrofolat (FH4) Perbedaannya dengan biosintesis purin pada terjadinya gugus ribosil=fosfat (ribosa-P), partisipasinau ribose –P ini agak lanjut. Dantahapan sebagai berikut: 1.Didahului oleh pembentukkan karbamoil-P, yang dihasilkan dari reaksi: antara glutamin, ATP, CO2 yang dikatalisis oleh enzim karbamoil sintase, dan reaksi ini berlangsung dimitokondria. 2. Karbamoil-P berkondensasi dengan asam aspartat menhasilkan karbamoil-aspartat, reaksi ini dikatalisis oleh enzim aspartat transkarbaoilase. 3. Dengan terbebaskan H2O maka terjadilah penutupan rantai karbamoil-aspartat menjadi dihidroorotat (DHOA = dihidroorotic acid), yang dikatalisis oleh enzim dihidro orotase. 4. Reaksi berikutnya pembentukan asam orotat (OA=orotic acid) dari DHOA yang dikatalisis oleh enzim DHOA dehidrogenase, koenzim NAD+.
Lanjutnya 5.Penambahan gugus ribosil fosfat pada asam orotat. Yang dikatalisis enzim orotao fosforibosiltransferase. Mengha silkan orotodilat (ortidin monofosfat =OMP). 6. Asam OMP mengalami dekarboksilasi dan menghasilakan uridilat atau (uridin monofospat = OMP). Yang reaksinya sebagai berikut. CO2 + Glutamin + ATP O O C H N CH C CH O N COO Aspartat transkarbamoilase O=C O CH CH H N COO 3 2 2 + H N-C-OP O 3 2 Dihidrootase Aspartat Asam karbamoil-aspartat CAA=Carbamoilaspartic acid Karbamoil-fosfat(CAP) O C HN CH O=C CH N COO H H O 2 2 Dihidrootat DH-ase Asam dihidroorotat DHOA=dihidroorotic acid Asam ortotat NAD NADH +H
O C HN CH O=C C - COOH N R-5P O C HN CH O=C C N R-5P Orotat fosforibosil transfrase O C HN CH O=C C - COOH N H Asam ortidilat dekarboksilase PRPP PPi Lanjutnya CO 2 UMP OMP Asam ortat ATP Ribonukleotida oksidase dUMP dUDP UDP deoksiuridindifosfat ATP N ,N FH4 Timidin sintetase H Folat ATP UTP O C HN C-CH O=C C N R-5P NH C HN CH O=C C N R-5P-P-P 2 Glutamin CTP sintetase 2 TMP CTP
Lintas penyelamatan pirimidin Primidin strukturnya lebih sederhana, lintas sintesisnya lebih cepat, dibandung purin, prazatnya hampir sama. (glutamin, glisisn, CO2 dan PPRibose fosfat). Sel pada hewan mamalia tidak mempunyai cara yang efisien untuk menyelematkan basa pirimidin, Senyawa ATP, ADP timidin dan TMP timidin kinase dapat menyelamatkan menjadi nukleotidanya masing-masing. Contoh: URIDIN UMP Enzim yang bertanggung jawab untuk biosintesis pirimidin adalah orotat fosforibosil transferase STIDIN CMP dapat menyelamatkan asam orotat menjadi TIMIDIN TMP IMP ATP ADP Uridinkinase ATP ADP Uridinkinase ATP ADP Timidinkinase
Lanjjutnya Tetapi diaggap tidak tepat karena asam orotat bukan basa pirimidin yang sempurna. Orotat fosfo ribosil transferase dapat menggaruhi pirimidin sebagai substratnya. Enzim tersebut juga mampu mengubah allopurinol (4-OH pirazolopirimidin) menjadi nukletida, dengan ribosil terikat pada N-1 cincin pirimidin. Enzim ini juga mampu merubah 5-flourourasil (obat anti kanker) dengan fosforilasi pada basa urasil. Sedangkan 2’-deoksisitidin dapat mengalami fosforilase oleh enzim deoksi sistidinkinase. Dan enzim ini juga dapat melakukan fosforilasi pada deoksiguanosin dan deoksiadenosin
Pengaturan biosintesis pirimidin Nukleotida purin PPRP + ATP +R5-P Aspartat ATP +CO2 +Glutamin CAP CAA DHO OA OMP UMP UDP UTP UDP Memacu + + Penghambatan CTP UDP dUDP TDP TMP
Katabolisme pirimidin Katabolisme pirimidin terjadi utamanya di hati, dan pembe basan CO2 merupakan jalur utama dari katabolisme urasil, sitosin dan timin dengan produk utama -alanin dan -aminoisobutirat. Dan timin adalah prekusor utama asam -aminoisobutirat. , O CH HN CH O NH NH N O NH 3 Timin Sitosin NADPH +H NADP 1/2 O NH O N O NH O CH HN CH O NH 3 3 H H dihidrotimin Urasil
Lanjutan NADPH +H NADP H O 2 O H HN C-H H O N H H Urasil dihidrotimin - OOC H N C - CH H C CH O N H OOC H N CH C CH O N H - H O 2 3 2 2 2 2 Asam uredobutirat b dihidrourasil Asam uredopropionat b H O 2 H O 2 + H NCH CH COO - 2 3 2 + - H NCH CH(CH )COO -alanin 3 2 3 b Asam aminoisobutirat CO + NH b 2 3 2 O CH COO 3 - Asam asetat
Beberapa Asam Amino Harus Diperoleh dari Makanan • Mahluk hidup berbeda dalam kemampuannya melakukan sintesis ke- 20 macam asam amino. Juga berbeda nitrogen yang dapat digunakan sebagai prekursor gugus amino. • Sebagai contoh, tikus albino dan manusia hanyadapat melakukan sintetis 10 dari 20 asam amino yang dibutuhkan sebagai unit pembangun bagi biosintesis protein. • Kesepuluh asam amino ini disebut asam amino nonesensial atau asam amino yang dapat diganti,yang dibentuk dari amonia dan berbagai sumber karbon. • Kesepuluh asam amino yang lain harus diperoleh dari makanan dan karenanya disebut nutritif esensial atau asam amino yang tidak dapat diganti. • Tumbuhan tingkat tinggi lebih beragam, dapat membuat semua asam amino yang dibutuhkan untuk sintesis protein. • Organisme tersebut dapat menggunakan baik amonia maupun nitrat sebagai prekursor gugus aminonya. Mikroorganismeberbeda-beda dalam kemampuannya melakukan sintesis asam amino.
Tabel Asam amino Nonesensial dan Esensial untuk Manusia dan Tikus Albino, *. Esensial untuk hewan dan manusis yang sedang tumbuh, sedangkan dewasa tidak
Sebagai contoh, Escherichia colidapat membuat semua asam amino yang dibutuhkan untuk sintesis protein dari prekursor sederhana, tetapi bakteri asam laktat tidak dapat dan harus memperoleh asam amino tertentu dari lingkungannya. • Glutamat, Ghitamin, dan Prolin Mengambil Bagian dalam Lintas Biosintetik Bersama Biosintesis asam amino nonesensial, yang dapat disintesis oleh manusia, tikus albino dan mamalia lainnya kebanyakan prekursor utama asam amino nonesensial adalah asam -ketonya, yang pada akhirnya ber-asal dari senyawa antara siklus asam sitrat. Gugus amino biasanya dilengkapi dengan reaksi transminasi dari glutamat, dikatalisis oleh transaminase, yang mengandung piridoksal fosfat sebagai gugus protestetik. • Gambar 1. Biosintesis golongan asam amino glutamat. Konversi • Glutamat menjadi prolin • Diketahui bahwa enzim pengikat glutamil 5-fosfat) adalah senyawa antara di dalam reaksi ini, yang terjadi dalam dua tahap. • Glutamat + ATP glutamil 5-fosfat + ADP Glutamil 5-fosfat + NH4+ glutamin + Pi + H+ • Jumlah: Glutamat + ATP + NH4+ glutamin + ADP + Pi + H+ • Reaksi di atas juga merupakan reaksi sentral yang penting di dalam metabolisme asam amino, karena merupakan lintas utama untuk mengubah amonia bebas, yang beracun, men-jadi glutamin tidak beracun untuk diangkut di dalam darah. Glutamin sintetase adalah enzim alosterik. Pada E. coli dan prokariota lainnya, aktivitas katalitik glutamin sintetase diatur oleh sejumlah metabolit yang berbeda, seperti yang nanti akan ditunjukkan. • Prolin, suatu senyawa turunan siklik glutamat, dibentuk oleh lintas yang ditunjukkan pada Gambar 2. Glutamat mula-mula direduksi menjadi senyawa --semialdehida, yang kemudian rnengalami siklisasi dan reduksi lebih lanjut menjadi prolin. • Glutamat kinase dehidrogenase H • OOC-CH-CH2CH2-COOH OOC-CH-CH2CH2-C=O • (Glutamat) (-semialdehid) • ATP+H- +NADPH ADP + NADP +Pi H2O (spontan) • COO- COO- • CH CH • NC CH2 H-N CH2 • Pirolin karboksilat reduktase • H2C CH2 HC CH2 • Prolin H+ +NaDPH NADP1-Prolin 5-karboksilat • Gambar 2.Ke lima atom karbon berasal dari asam glutamat. Protin adalah suatu penghambat alosterik pada reaksi awal biosintesisnya. Aktivitas umpan balik negatif ini ditun-jukkan oleh tanda panah dan titik peng-hambatan oleh garis berwarna terputus-putus • Alanin, Aspartat dan Asparagin Juga Berasal dari Metabolit Sentral • Pada kebanyakan organisme, asam amino nonesensial alanindan aspartatberturut-turut berasal dari piruvat dan oksaloasetat oleh transaminasi dari glutamat. • Glutamat + piruvat -ketoglutarat + alanin • Glutamat + oksaloasetat -ketoglutarat + aspartat • Dalam banyak bakteri, aspartat adalah prekursor langsung asparagin dalam suatu reaksi yang dikatalisis oleh asparagin sintetase,analog dengan reaksi glutamin sintetase. • Aspartat + NH4+ + ATP asparagin + ADP + Pi + H+ • -OOC—CH—CH2—COO- Aspartat • +NH3 • + -OOC—CH—CH2—CH2 C=O Glutamin • +NH3 NH2 • ATP • AMP+PPi asparagin sintetase • OOC—CH—CH,—C=O Asparagin • +NH3 NH2 • -OOC—CH,— CH2-CH2 COO" Glutamat • +NH3 • Gambar 3. Biosintesis asparagin dan glutamin adanya aktivitas enzim aparagin • sintetase • Tetapi, pada mamalia ada perbedaan lintas untuk sintesis asparagin: gugus amino dipindahkan dari gugus amida glutamin menjadi gugus -karboksil aspartat (Gambar 22-3) oleh asparagin sintetase(bergantung ATP). • Glutamin + aspartat + ATP + H2 O glutamat + asparagin + AMP + PPi • Tirosin Dibuat dari Asam Amino Esensial, Fenilalanin • Tirosinadalah asam amino nonesensial, tetapi dibuat oleh hewan dari asam amino esensial fenilaninmelalui hidroksilasi pada posisi 4 pada gugus fenil oleh fenilanin oksigenase, yang juga ikut serta dalam degradasi fenilalanin. Dalam reaksi ini NADPH dibutuhkan sebagai kopereduksi molekul oksigen; ingat kembali bahwa fenilalanin oksigenase adalah suatu monooksigenaseatau oksidase berfungsi ganda. Reaksi yang dikatalisis adalah: • Fenilalanin + NADPH + H+ + O2 tirosin + NADP+ + H2 O. • -CH2-CH- COOH HO- -CH2-CH-COOH • NH2 NH2 • Sistein Dibuat dari Dua Asam Amino Lainnya, Metionin dan Serin • Pada mamalia, sisteindibuat dari dua asam amino lainnya, metionin yang esensial di dalam makanan dan serinyang tidak esensial. Metionin memberikan atom sulfur dan serin memberikan kerangka karbon pada sintesis sistein. Pada reaksi pertama di dalam lintas ini, metionin diubah menjadi S-adenosilmetionm(Gambar 4) oleh reaksi dengan ATP.
Lintas biosintesis menuju hubungan asam amino glutamat, glutamin dan prolin (Slide 26) adalah sederhana dan nampaknya identik dalam semua bentuk kehidupan. Glutamatdibentuk dari amonia dan -ketoglutarat, suatu senyawa antara siklus asam sitrat, melalui kerja L-glutamat dehidrogenase.Tenaga pereduksi yang diperlukan diberikan oleh NADPH. NH4+ + -ketoglutarat + NADPH L-glutamat + NADP+ + H2O Reaksi ini adalah dasar yang penting di dalam biosintesis semua asam amino, karena glutamat adalah donor gugus amino dalam biosintesis asam amino yang lain melalui reaksi transaminasi L-glutamat dehidrogenase terletak di dalam matriks mitokondrion. Glutamin.dibentuk dari glutamat melalui kerja glutamin sintetase.
Glutamat + NH4+ +ATP glutamin + ADP + Pi + H+ O Glut.dehidrogenase NH2 HOOC-C-CH2CH2-COOH HOOC-C-CH2CH2 (-ketoglutarat)HOOC H +NADPH+NH3NADP ATP +NH4+ `ADP + Pi + H+ ` COOH H2N-C-H CH2 CH2 C=O NH2 COOH C HN CH HC CH Spontan Prolin Glutamin
Diketahui bahwa enzim pengikat glutamil 5-fosfat) adalah senyawa antarara di dalam reaksi ini, yang terjadi dalam dua tahap. • 1.Glutamat + ATP glutamil 5-fosfat + ADP • 2.Glutamil 5-fosfat + NH4+ glutamin + Pi + H+ • Jumlah: Glutamat + ATP + NH4+ glutamin +ADP+Pi+ H+ • Reaksi di atas juga merupakan reaksi sentral yang penting di dalam metabolisme asam amino, karena merupakan lintas utama untuk mengubah amonia bebas, yang beracun, men-jadi glutamin tidak beracun untuk diangkut di dalam darah. • Glutamin sintetase adalah enzim alosterik. Pada E. coli dan prokariota lainnya, aktivitas katalitik glutamin sintetase diatur oleh sejumlah metabolit yang berbeda, seperti yang nanti akan ditunjukkan.
Prolin, suatu senyawa turunan siklik glutamat, dibentuk oleh lintas yang ditunjukkan pada slide 15. Glutamat mula-mula direduksi menjadi senyawa --semialdehida, yang kemudian rnengalami siklisasi dan reduksi lebih lanjut menjadi prolin. Glutamat kinase dehidrogenase H OOC-CH-CH2CH2-COOH HOOC-CH-CH2CH2-C=O (Glutamat) (-semialdehid) ATP+H- +NADPH ADP + NADP +Pi H2O(spontan) COO- COO- CHCH HN CH2H-N CH2 Pirolin karboksilat reduktase H2C CH2 HC CH2 Prolin1-Prolin 5-karboksilat H+ +NaDPH NADP Ke lima atom Prolin karbon berasal dari asam glutamat. .
Prolin adalah suatu penghambat alosterik pada reaksi awal biosintesisnya. Aktivitas umpan balik negatif ini ditunjukkan oleh tanda panah dan titik peng-hambatan oleh garis berwarna terputus-putus • Alanin, Aspartat dan Asparagin Juga Berasal dari Metabolit Sentral • Pada kebanyakan organisme, asam amino nonesensial alanindan aspartatberturut-turut berasal dari piruvat dan oksaloasetat oleh transaminasi dari glutamat. • Glutamat + piruvat -ketoglutarat + alanin • Glutamat + oksaloasetat -ketoglutarat + aspartat • Dalam banyak bakteri, aspartat adalah prekursor langsung asparagindalam suatu reaksi yang dikatalisis oleh asparagin sintetase,analog dengan reaksi glutamin sintetase. • Aspartat + NH4+ + ATP asparagin + ADP + Pi + H+
--OOC—CH—CH2—COO- Aspartat +NH3 + -OOC—CH—CH2—CH2 C=O Glutamin +NH3 NH2 ATP AMP+PPi asparagin sintetase OOC—CH—CH,—C=O Asparagin +NH3 NH2 -OOC—CH,— CH2-CH2 COO" Glutamat +NH3 Biosintesis asparagin dan glutamin adanya aktivitas enzim aparaginsintetase • Tetapi, pada mamalia ada perbedaan lintas untuk sintesis asparagin: gugus amino dipindahkan dari gugus amida glutamin menjadi gugus -karboksil aspartat (Gambar 22-3) oleh asparagin sintetase(bergantung ATP). • Glutamin + aspartat + ATP + H2 O glutamat + asparagin + AMP + PPi
Tirosin Dibuat dari Asam Amino Esensial, Fenilalanin • Tirosinadalah asam amino nonesensial, tetapi dibuat oleh hewan dari asam amino esensial fenilaninmelalui hidroksilasi pada posisi 4 pada gugus fenil oleh fenilanin oksigenase, • Enzim ini juga ikut serta dalam degradasi fenilalanin. Dalam reaksi ini NADPH dibutuhkan sebagai kopereduksi molekul oksigen; ingat kembali bahwa fenilalanin oksigenase adalah suatu monooksigenaseatau oksidase berfungsi ganda. Reaksi yang dikatalisis adalah: Fenilalanin + NADPH + H+ + O2 tirosin + NADP+ + H2 O. -CH2-CH- COOHHO- -CH2-CH-COOH NH2NH2 • Hasil akhir rangkaian reaksi yang kompleks ini adalah penggantian gugus —OH pada serin dengan suatu gugus SH, yang berasal dari metionin, untuk membentuk sistein (Gambar 5). • COO- COO- • H2N-C-H H2N-C-H Sistein • CH2 metionin CH2 • CH2 SH • S • CH3-Ketoglutarat Sistationin • H2O + ATP + NH4+-liase • Metionin adenil transferase • PPi + Pi • COO- COO- • H2N- C- H H2N-C-H • CH2 S-Adenometilmetionin CH2 • CH2 CH2 • S CH2 O Adenin S – CH2 – CH-COO- • CH3 H H SH • H H • Penerima gg metil OH OH H2O sistationin • -sintetase • Senyawa termetilasi Reaksi pemindahan metil Serin • COO- COO- • H2N- C- H H2N-C-H • CH2 S-Adenosilhomosestein CH2 • CH2 H2O Adenin CH2 • S CH2 O Adenin SH • H H Adenosilhomosistein • H H sintetase. Homosistein • OH OH
L-Metionin + ATP + H2O S-adenosilmetionil + PPi- + Pi NH2 COO N N H2N- C- H CH Adenin CH2N N CH2 S CH2 O Ribose CH3 H H H H • Metionin OH OH Struktur S-adenosilmetionin. Atom S stabil pada ikatan tertier.
Gugus adenosil dapat dianggap sebagai pembawa molekul metionin. Dalam bentuk ini, gugus metil pada metionin bersifat sangat reaktif dan dapat dipindahkan secara enzimatik ke berbagai senyawa penerima gugus metil, meninggalkan S-adenosilhomosisteinsebagai produk dimetilasi. S- Adenosilmetionin + penerima gugus metil • S-adenosilhomosistein + penerima termetilasi Sebagai contoh, telah diketahui bahwa S-adenosil metionin berperan sebagai donor gugus metil di dalam pengubahan fosfatidiletanolamin menjadi fosfatidilkolin. • Setelah penarikan gugus metil, S-adenosilhomosistein siap untuk memasuki reaksi selanjutnya pada lintas menuju sistein, seperti diperlihatkan pada Pada tahap selanjutnya, homosistein bebas dibentuk oleh reaksi. S-Adenosilhomosistein + H2 O adenosin + homosistein
Homosistein lalu bereaksi dengan serin dalam reaksi yang dikatalisis oleh sistationin -sintetase menehasilkan sistationin • Homosistein + serin sistationin + H2O • Pada tahap terakhir, sistationin S-liaseyang juga merupakan enzim piridoksal fosfat, mengkatalisis pemindahan amonia dan membelah sistationia, menghasilkan sistein bebas • Sistationin + H+-ketobutirat + NH4+ + sistein • Bila digabungkan semua tahap-tahap yang terpisah tadi bersama-sama, peramaan keseluruhan bagi sintesis sistein menjadi: L-Metionin+ ATP + penerima metil + H2O + H+ + serin penerima termetilasi + adenosin + -asetobutirat + NH4+ + sistein + PPi + Pi